Hệ vận động
Gs. Bùi Tấn Anh - Võ Văn Bé - Phạm Thị Nga
![]()
| 1- Hệ xương và cơ của động vật có xương sống |
| 2- Các loại cơ |
| 1- Ðặc điểm chung của sự co cơ xương |
| 2- Cơ sở phân tử của sự co cơ |
| 3- Kiểm soát điện hóa của sự co cơ |
|
Các cơ quan hiệu ứng là một
bộ phận của sinh vật giúp cho chúng
đáp ứng với các kích thích.
Hoạt động của các cơ quan
nầy rất khác nhau như sự tiết
của tuyến, sự tạo ra ánh sáng của
đom đóm, sự địa hướng
động và quang hướng động
của thực vật, chuyển động
của tế bào chất trong tế bào, và quen
thuộc nhất là các cử động của
cơ ở động vật.
Như chúng ta đã thấy ở phần
I, thực vật cũng có khả năng cử
động chậm để đáp ứng
với ánh sáng và trọng lực.
Các cử động được
tạo ra do sự tăng trưởng khác nhau
được kiểm soát bởi hormon.
Nhiều thực vật cũng có khả năng
cử động nhanh như lá cây Trinh nữ
(Mimosa pudica) khép lại chỉ một vài giây sau
khi bị chạm vào.
Như vậy rõ ràng là hoạt động
chuyển động không phải là một
đặc tính riêng của động vật.
Tuy nhiên, một điều vẫn đúng
là phần lớn các chi tiết về sự
tạo ra các chuyển động được
tìm thấy ở giới động vật và
đó cũng là chủ đề chính mà chúng
ta sẽ tập trung ở chương nầy.
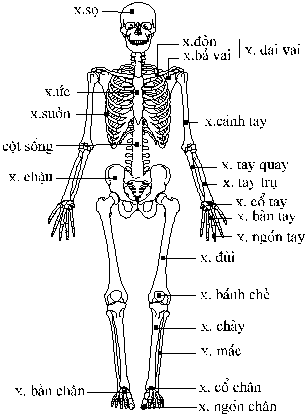
Bộ
xương của động vật có xương
sống được tạo thành chủ
yếu từ xương hoặc sụn.
Sụn là một mô liên kết mà trong
đó chất dịch nền gian bào có các
sợi protein rất mãnh.
Nó chắc chắn nhưng không cứng và
giòn như xương.
Trong tất cả các phôi của động
vật có xương sống, bộ xương
được tạo thành từ sụn.
Bộ xương sụn được duy
trì ở cơ thể trưởng thành của
một vài dạng như cá mập, cá đuối.
Tuy nhiên ở phần lớn động
vật có xương sống, trong quá trình phát
triển sụn dần dần được
thay thế bằng xương và chúng chỉ còn
lại ở những vùng cần có sự
phối hợp giữa độ chắc và
độ mềm dẽo như là ở đầu
xương sườn, ở bề mặt xương
tại các khớp, trong thành của thanh quản
và khí quản, ở tai ngoài và ở mũi. Xương
gồm một chất cơ bản bằng
protein, có nhiều ion vô cơ đặc biệt
là phosphat calci.
Xương là một mô sống, nó có các
tế bào và được cung cấp nhiều
mạch máu. Có
hai loại xương cơ bản là xương
xốp và xương đặc.
Xương xốp (spongy bone) được
tìm thấy ở phần trung tâm của xương,
gồm một mạng lưới của các
thỏi cứng, khoảng giữa chúng được
lắp đầy chất dịch.
Phía ngoài của xương có các xương
đặc (compact bone), những phần cứng
của chúng xuất hiện như một
khối liên tục và chỉ có những xoang
rất nhỏ. Xương
đặc được hợp thành từ
những đơn vị cấu trúc gọi là
hệ Havers (Haversian system) chạy dọc suốt
chiều dài của xương.
Mỗi đơn vị có hình trụ và
được hợp thành từ các lớp
chất nền có chứa Ca sắp xếp đồng
tâm quanh một ống Havers (Haversian canal) ở
trung tâm. Các
mạch máu và các dây thần kinh đi qua
những ống nầy.
Các tế bào xương nằm trong các
xoang nhỏ trong chất nền gian bào và
được nối bởi một hệ
thống các ống cực nhỏ (canaliculi) xuyên
ngang qua các lớp chất nền.
Sự trao đổi chất giữa các
tế bào xương và các mạch máu trong
ống Havers diễn ra trong những ống
cực nhỏ nầy (Hình 1).
Hình
1. Ảnh
một lát cắt ngang qua xương Bộ
xương của động vật có xương
sống thường được chia làm hai
phần: (1) xương
trục (axial skeleton) là phần trục chính
dọc theo chiều dài cơ thể, bao gồm
hộp sọ, cột sống và các xương
sườn; (2) xương chi (appendicular skeleton)
bao gồm các xương nối với trục
như xương chi, xương đai và xương
chậu. (Hình
2).
Hình
2. Bộ xương
của người Một
số xương được nối bằng
các khớp bất động (suture) như
một số xương nhỏ hợp thành xương
sọ. Một
số khác được giữ với nhau
tại các khớp động nhờ các dây
chằng (ligament). Các
cơ xương nối với xương
nhờ các gân (tendons) giúp cho các xương có
thể uốn gập lại tại các khớp.
Lực gây ra cử động nầy luôn
luôn là lực kéo do sự co cơ.
Sự duỗi thẳng hay chuyển hướng
của các xương tại khớp này
phải được hoàn tất nhờ sự
co của các bộ cơ có chức năng đối
kháng.
Nếu một cơ gắn với hai xương
có một hoặc nhiều khớp thì sự co cơ
chỉ làm cho một xương chuyển động.
Một cơ đôi khi có nhiều điểm
gắn trên cùng một xương hoặc trên
nhiều xương khác nhau.
Các hoạt động được
tạo ra từ sự co cơ phụ thuộc vào
vị trí của những điểm gắn
nầy và vào loại khớp giữa các xương.
Thật vậy, trong trường hợp bình
thường, hệ thần kinh chỉ truyền
xung động đến các cơ hoạt động
trong một nhóm cơ đối kháng.
Vì vậy khi một cơ này co thì cơ
đối lập sẽ duỗi ra, giúp cho xương
chuyển động.
Ngoài ra còn có các cơ điều phối
(synergists) giữ vai trò hướng dẫn và
giới hạn các cử động.
Ở
động vật có xương sống người
ta thường phân biệt ba loại cơ chính
là cơ xương, cơ trơn và cơ tim
(Bảng 1)
a.
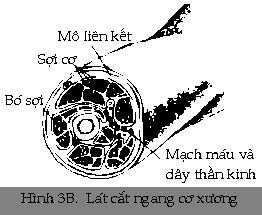
Cơ xương (skeletal muscle)
sợi
thường được bọc bởi các mô
liên kết thành các bó, các bó này lại
được bọc chung với nhau bởi
nhiều mô liên kết tạo thành cơ.
Như vậy, một cơ được
hợp thành bởi nhiều bó sợi cơ,
giống như một dây thần kinh được
hợp thành từ nhiều bó sợi thần
kinh (Hình3B).
Có ít nhất hai loại cơ xương:
cơ đỏ và cơ trắng.
Cơ đỏ rất giàu mạch máu,
nhiều
ty thể và myoglobin. Myoglobin giống như
Hemoglobin có khả năng kết hợp lõng
lẽo với oxy và dự trử chúng trong cơ.
Cơ đỏ oxy hóa các acid béo như là
nguồn năng lượng chính. Mặc dù
chúng co tương đối chậm nhưng có
khả năng hoạt động trong một
thời gian dài mà không bị mõi.
Ngược lại, cơ trắng có ít
mạch máu, ít ti thể hơn và lượng
myoglobin thấp. Nó
hầu như phụ thuộc vào sự phân
giải yếm khí của glycogen để cung
cấp năng lượng và có khả năng
co nhanh, mạnh nhưng chỉ trong một
thời gian ngắn vì mõi cơ nhanh chóng. b.
Cơ trơn (smooth muscle)
Nhiều sự khác biệt trong chức năng
giữa cơ xương (đáp ứng với
những thay đổi của môi trường
ngoài) và cơ trơn (đáp ứng với
những thay đổi của môi trường
trong) được phản ánh trong mối liên
hệ khác nhau với hệ thần kinh.
Các tế bào của cơ xương
được phân bố chỉ một dây
thần kinh từ hệ thần kinh dinh dưỡng:
chúng co khi bị kích thích bởi xung thần kinh
và duỗi khi không có xung kích thích.
Ngược lại, các tế bào cơ trơn
được phân bố bởi hai dây thần
kinh; một từ hệ thần kinh giao cảm
và một từ hệ thần kinh phó giao
cảm. Chúng co
lại để đáp ứng với xung
thần kinh từ một sợi và bị ức
chế (không co) bởi xung đến từ
một sợi khác. Cơ
xương không thể hoạt động bình
thường nếu thiếu liên hệ thần
kinh và sẽ bị thoái hóa nhưng cơ trơn
có thể co mà không cần kích thích thần kinh
như trường hợp nhu động
ruột chẳng hạn. c.
Cơ tim (cardiac muscle)
II.
SINH LÝ HỌC CỦA HOẠT ÐỘNG CƠ
Các
sợi cơ và các tế bào thần kinh
giống nhau ở chỗ cả hai sẽ đáp
ứng chỉ khi một kích thích đạt
đến ngưỡng cường độ
và thời gian. Giống
như tế bào thần kinh, sợi cơ cũng
có thuộc tính tất cả hoặc không (all or
none). Nếu như
một sợi cơ tách rời chịu một
kích thích có giá trị trên ngưỡng, độ
co cơ nhận được đều
giống nhau, bất kể giá trị nào của
kích thích. Mặc
dù một sợi cơ đáp ứng theo
kiểu tất cả hoặc không nhưng
một cơ được hợp thành bởi
nhiều sợi cơ thì lại khác. Có thể
dễ dàng chứng minh trong phòng thí nghiệm
rằng cơ có khả năng đáp ứng
ở những mức độ khác nhau phụ
thuộc vào cường độ kích thích:
nếu một cơ tiếp nhận một kích
thích ở cường độ ngưỡng, cơ
sẽ co rất yếu.
Khi cường độ kích thích mạnh
hơn được tạo ra sau một ít giây
nghỉ, cơ sẽ co hơi mạnh hơn.
Cường độ càng tăng làm cơ
co càng mạnh cho đến khi sự gia tăng
kích thích không làm tăng cường độ
của đáp ứng. Lúc này cơ đã đạt
đến đáp ứng tối đa. Làm thế nào có thể giải thích các kết quả nầy nếu các sợi cơ đáp ứng theo nguyên lý tất cả hoặc không? Ðó là do sự tương tác giữa các sợi cơ khác nhau trong mỗi cơ. Vì trong một cơ, mỗi sợi cơ có giá trị ngưỡng khác nhau, được phân bố bởi các sợi thần kinh khác nhau và những sợi này không phải đều bị kích thích đồng thời, do đó mặc dù từng sợi cơ đáp ứng với kích thích theo kiểu tất cả hoặc không nhưng một sự gia tăng cường độ của kích thích trên mức độ ngưỡng có thể dẫn tới một đáp ứng lớn hơn của cơ do nhiều sợi cơ bị kích thích. Tuy nhiên cơ sẽ đạt đến mức đáp ứng tối đa khi toàn bộ sợi cơ bị kích thích và sự gia tăng cường độ hơn nữa cũng không gây ra sự đáp ứng (Hình 6).
Hình
6. Ðáp ứng
của một cơ với những kích thích có
cường độ khác nhau
Thí
nghiệm với cơ Êúch cho thấy nếu
một kích thích thích hợp tác động lên
cơ, sẽ có một thời gian ngắn trong
đó sự co cơ không xảy ra.
Ðây là thời kỳ tiềm phục
(latnt period) thường thay đổi từ
0,0025 đến 0,004 giây.
Tiếp theo là thời kỳ co
(contraction period) và ngay sau đó là thời
kỳ duỗi (relaxation period).
Ba thời kỳ nầy tạo thành một
co cơ đơn (simple twich) (Hình 7).
Mỗi
co cơ đơn cần có một thời gian
nghỉ thích hợp giữa hai kích thích kế
tiếp. Nếu
có một loạt kích thích liên tiếp tác động
lên cơ, cơ chưa kịp duỗi hoàn toàn
khi đáp ứng với kích thích trước thì
kích thích kế tiếp đã đến.
Trong trường hợp này biên độ
co cơ sẽ lớn hơn co đơn độc,
gọi là sự cộng co (summation).
Khi các kích thích lặp lại rất nhanh,
cơ không thể duỗi giữa các lần kích
thích. Trong trường
hợp nầy không thể phân biệt được
từng co cơ đơn, chúng hợp lại thành
co cứng (tetanus) (Hình 8).
Nếu cơ co cứng quá lâu, chúng sẽ
bị mõi và biên độ co cơ sẽ
giảm dần ngay cả
khi kích thích liên tục với cùng cường
độ. Sự
mõi cơ có liên quan đến việc giảm lượng
glycogen tích trử, sự tích tụ acid lactic và
những thay đổi hóa học khác.
A
B Hình 9. Co đẳng trương (A) và co đẳng trường (B)
a.
Năng lượng cho sự co cơ
Năng
lượng cần cho sự co cơ đến
từ ATP. Chất
nầy lại được tạo ra từ
sự biến dưỡng của glucoz và acid béo.
Nhưng có ít
ATP được dự trử trong cơ đến
nỗi chỉ một vài lần co cơ sẽ
nhanh chóngbị mất đi nguồn cung cấp
năng lượng. Như
vậy cơ vượt qua hạn chế này như
thế nào? Mặc dù có ít ATP nhưng lại có
một hợp chất khác có nhóm phosphat
được dự trử phổ biến trong
cơ là creatin phosphat.
Chất này được tạo thành
bằng sự gắn một nhóm phosphat vào
chất creatin. Creatin
phosphat không được sử dụng
trực tiếp để cung cấp năng lượng
cho sự co cơ nhưng nó có thể chuyển
nhóm phosphat đến ADP để thành lập
ATP:
Creatin Phosphat + ADP + H+ ---> Creatin + ATP Chất
ATP mới được thành lập tác động
như nguồn năng lượng trực
tiếp cho sự co cơ.
Cơ dự trử đủ creatin phosphat
để có thể co trong nhiều giây trước
khi sự hô hấp tế bào có thể sản
xuất thêm ATP.
Nếu yêu cầu của cơ không
lớn, năng lượng bổ sung có thể
đến từ sự oxy hóa hoàn toàn của
glucoz hoặc acid béo thành CO2 và H2O với sự
tham gia của O2. Trong
thời gian nghỉ trước khi hệ hô
hấp và hệ tuần hoàn tăng lượng
oxy cung cấp cho hoạt động của cơ,
một số oxy cần cho sự hô hấp
hiếu khí trong cơ đỏ được
cung cấp bởi myoglobin. Trong
trường hợp cơ phải hoạt động
tận lực, yêu cầu về năng lượng
của cơ (nhất là cơ trắng) rất
lớn và oxy từ myoglobin nhanh chóng được
sử dụng hết.
Vì oxy không được các mô thu nhận
kịp thời nên cơ phải nhận năng
lượng cần thiết từ quá trình hô
hấp yếm khí bằng sự lên men lactic và
tạo ra món nợ oxy (oxygen debt).
Một số ít acid lactic tích tụ trong cơ
còn phần lớn khuếch tán vào các mao
mạch của cơ và được máu chuyên
chở về gan. Khi
hoạt động nặng chấm dứt,
sự thở sâu hoặc thở gấp sẽ
cung cấp cho gan một lượng lớn oxy
cần cho sự hô hấp hiếu khí, vì
vậy trả được món nợ oxy.
Trong gan, acid lactic được biến
đổi thành acid pyruvic mà hầu hết
sẽ bị oxy hóa để tạo ra ATP.
Acid lactic còn lại được dùng
để tổng hợp glucoz và glycogen. b.
Cơ chế co cơ
Như
ta đã biết, một cơ xương
được đặc trưng bởi các vân
và được hợp thành bởi một
số các sợi cơ.
Khảo sát những sợi này với độ
phóng đại rất cao cho thấy chúng
được tạo thành từ nhiều
đơn vị cấu trúc dài và mãnh gọi là
tơ cơ (myofibril).
Mỗi tơ cơ có đường kính
khoảng 1-2 (m, có các ty thể nằm trong
tế bào chất giữa chúng.
Các vân được hình thành do sự xen
kẻ giữa các đĩa sáng (đĩa I) và
các đĩa tối (đĩa A).
Giữa mỗi đĩa A có một vùng sáng
hơn gọi là vùng H.
Giữa mỗi đĩa I có một
vạch mãnh, rất tối gọi là vạch Z.
Toàn bộ một vùng từ vạch Z này
đến vạch Z kế tiếp gọi là
một sarcomer. Sarcomer
là đơn vị chức năng của sự
co cơ (Hình 10). Hình
10. Ảnh hiển vi cơ xương của
thỏ Các
vân của cơ xương phản ánh về
cấu trúc của các đơn vị chức năng
trong sự co cơ. Khi
sợi cơ co, sarcomer trở nên ngắn hơn
và độ rộng tương đối
của các đĩa thay đổi: đĩa I
và vùng H hẹp hơn, các đĩa A ít thay
đổi nhưng di chuyển đến gần
nhau hơn. Những
nghiên cứu của cơ dưới kính
hiển vi điện tử cho thấy mỗi tơ
cơ lại gồm có hai loại sợi
(filament), một sợi dầy và một sợi
mỏng, sắp xếp theo một kiểu
nhất định. Các
sợi dầy được tìm thấy trong
đĩa A còn các sợi mỏng chủ yếu
có trong đĩa I nhưng kéo dài ra đến
đĩa A. Sự
phân bố này giải thích được vì sao
có sự hiện diện của đĩa A,
đĩa I và vùng H.
Mỗi đĩa A (tối) là chiều dài
của vùng có các sợi dầy, tối nhất
ở vùng ranh giới nơi các sợi dầy và
mỏng chồng lên nhau, vùng H là nơi chỉ có
các sợi dầy.
Mỗi đĩa I tương ứng
với một vùng chỉ có các sợi mỏng.
Vạch Z là một cấu trúc hình
đĩa gắn vào các sợi mỏng, giữ
cho chúng ở vị trí thích hợp.
Các phân tích hóa học cho thấy sợi
dầy được hợp thành từ protein là
myosin còn những sợi mỏng chủ yếu là
protein actin. Những
quan sát này dẫn đến một học
thuyết về sự co cơ : các tơ cơ
lồng vào nhau do các sợi trượt lên
nhau. Vùng
chồng lên nhau giữa sợi dày và sợi
mỏng sẽ tăng lên cho đến khi các
sợi mỏng từ đĩa I ở hai phía
của một đĩa A gặp nhau.
Sự trượt này làm giảm chiều
rộng của vùng H, thậm chí làm mất hoàn
toàn vùng nầy (khi các sợi mỏng gặp
nhau). Chuyển
động của các sợi cũng kéo các
vạch Z lại gần nhau và làm giảm nhanh
chiều rộng của đĩa I.
Chiều rộng của đĩa A chỉ
thay đổi một ít vì vùng này tương
ứng với các sợi dày còn chiều dài
hầu như không đổi (Hình 12).
Như vậy học thuyết về sự
trượt của các sợi (sliding-filament
theory) giải thích được những thay
đổi có thể thấy trong sarcomer.
Tuy nhiên, vẫn còn phải trả lời
cho câu hỏi làm thế nào sự trượt
xảy ra. Hình
15. Sarcomere
với lưới cơ tương và hệ
thống T
Các ống của hệ thống T ăn sâu
vào trong màng tế bào cho phép điện thế
động lan truyền qua bề mặt tế bào
vào bên trong sợi cơ.
Ðiện thế động di chuyển nhanh
hơn sự khuếch tán của các ion, đủ
để các kích thích đi đến tất
cả các tơ cơ vì vậy các tơ cơ
ở gần bề mặt và những tơ co
ở trung tâm có thể co cùng một lúc.
Mối quan hệ giữa hệ thống T và
lưới cơ tương cho thấy rằng
một điện thế động lan
truyền dọc theo màng của hệ thống T
có thể làm thay đổi thuộc tính màng
của lưới cơ tương kế
cận. Ðiều
này rất quan trọng vì lưới cơ tương
có một lượng rất lớn ion Ion
Khi
cơ ở trạng thái nghỉ protein điều
hòa này ngăn cản sợi actin gắn vào
đầu myosin bằng cách khóa vị trí
gắn của myosin trên phân tử actin.
Phức hệ troponin cũng có hình cầu
và xuất hiện ở dạng bộ ba.
Mỗi phức hệ có ba vị trí liên
kết: một cho actin, một cho tropomyosin và
một cho ion Như
vậy chúng ta đã có đầy đủ các
thành phần của mô hình về hoạt động
của cầu nối và sự kích thích của
nó. Trong một
cơ nghỉ, cầu nối (đầu myosin
của sợi dầy) đã được
hoạt hóa bởi ATP nhưng chưa gắn
với sợi mỏng vì tropomyosin đã khóa
vị trí gắn phân
tử actin (Hình 17A).
Khi kích thích từ một tế bào
thần kinh vận động làm phát sinh
một điện thế động, điện
thế động sẽ lan truyền dọc theo
bề mặt cơ và đi vào các ống T,
từ đó kích thích lưới cơ tương
phóng thích ion Hình
17. Mô hình
về họat động của
thể gắn vào actin (Hình 17B), sau đó chúng
gập
các cầu nối trong
sự co cơ
lại kéo sợi actin về phía chúng, làm
cho các sợi trượt dọc lên nhau (Hình17C).
Khi một phân tử ATP mới được
gắn vào đầu myosin, đầu sẽ tách
ra khỏi sợi actin (Hình 17D).
Sau đó ATP bị thủy phân thành ADP và
Pi làm cho đầu bị đẩy ngược
trở lại vị trí ban đầu (Hình 17E).
Khi có nhiều ion
Có thể tóm tắt các bước trong quá
trình co cơ như sau: 1.
Xung thần kinh gây ra sự phóng thích
acetylcholin tại diện tiếp hợp thần
kinh- cơ 2.
Acetylcholin gắn vào thể tiếp nhận
trên màng sợi cơ, mở cổng ion.
Ion chạy qua màng
tế bào, khử phân cực chúng và tạo ra
điện thế động. 3.
Ðiện thế động lan truyền
dọc theo bề mặt tế bào cơ và đi
vào ống T. 4.
Ðiện thế động trong ống T làm
thay đổi tính thấm của màng lưới
cơ tương gây ra
sự phóng thích ion Ca dự trử. 5.
Ion Ca gắn vào phức hệ tropomyosin làm
thay đổi vị trí của chúng, không còn
che phủ vị trí gắn myosin trên actin. 6.
Ðầu myosin được hoạt hóa
bởi sự thủy phân ATP sẽ gắn vào
actin, ADP và Pi được phóng thích, đầu
myosin gập lại kéo sợi actin về hướng
myosin. 7.
ATP mới được gắn vào đầu
myosin, sau đó tách chúng ra khỏi actin.
ATP bị thủy phân làm cho đầu quay
trở lại vị trí cũ. 8.
ADP
và Pi vẫn gắn trên đầu myosin giúp chúng
có thể liên kết một lần nữa
với actin.
|