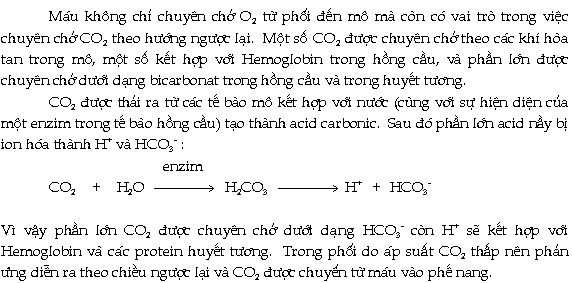
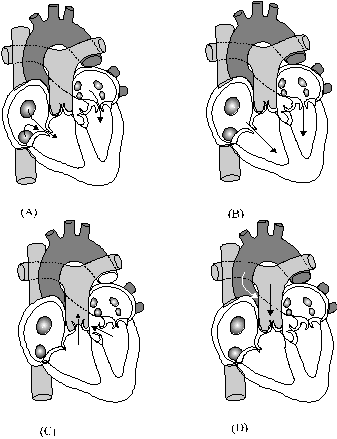Máu, hệ tuần hoàn và đáp ứng miễn nhiễm ở động vật hữu nhũ
Gs. Bùi Tấn Anh - Võ Văn Bé - Phạm Thị Nga
![]()
|
CHƯƠNG
V MÁU,
HỆ TUẦN HOÀN VÀ ÐÁP ỨNG MIỄN
NHIỄM
Ở
ÐỘNG VẬT HỮU NHŨ
Mọi tế bào sống trong cơ thể
động vật đều cần được
cung cấp các chất dinh dưỡng và O2 đồng
thờiphải loại bỏ các chất thải như CO2 và
các hợp chất có N2 .
Ở một số động vật,
mỗi tế bào đều tiếp xúc trực
tiếp hoặc được bao trong một môi
trường từ đó chúng có thể thu
nhận các chất cần thiết và loại
bỏ các chất thải.
Những động vật lớn hơn,
cấu trúc phức tạp hơn có hàng tỉ
tế bào nằm rất xa bề mặt cơ
thể và môi trường chung quanh.
Chúng cần phải có một hệ chuyên
chở đặc biệt bên trong cơ thể
(hệ tuần hoàn) để phân phối các
chất đến từng tế bào và thải
các chất bả.
Như
chúng ta đã biết máu là một loại mô liên
kết với một chất dịch cơ
bản và các yếu tố hữu hình.
Chất dịch cơ bản của máu
được gọi là huyết tương
(plasma). Lơ
lững trong huyết tương là các thành
phần tế bào, gồm 3 loại chính là: (1) Các
hồng cầu (erythrocytes); (2) các bạch cầu
(leukocytes) và (3) các tiểu cầu hay tấm máu
(platelets). Cả
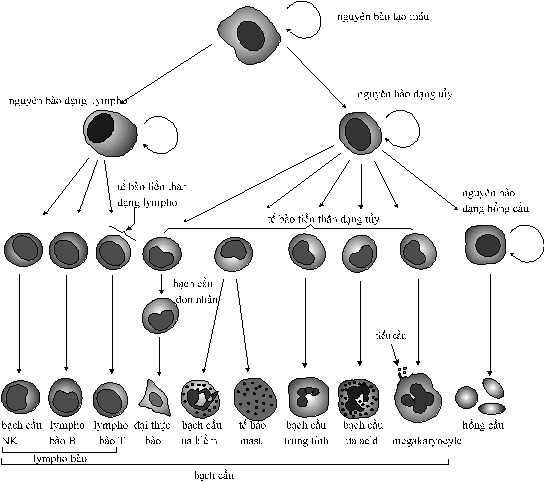
ba loại tế bào nầy đều xuất phát
từ các tế bào mô liên kết đặc
biệt gọi là các nguyên bào (stem cell) trong
tủy xương của cá thể trưởng
thành (Hình 1). Nếu
toàn bộ máu được xử lý để
chống đông và cho vào một ống
nghiệm đặt thẳng đứng, các
yếu tố tế bào sẽ từ từ
lắng xuống đáy, để lại
huyết tương ở phía trên.
Bình thường các tế bào chiếm
khoảng 40 - 50% tổng thể tích của máu,
trong khi huyết tương chiếm khoảng 50
- 60%. a.
Huyết tương Thành
phần cơ bản của huyết tương
là nước, chiếm khoảng 90%.
Trong nước có một số lượng
rất lớn các châtú hòa tan; nồng độ
của các chất nầy thay đổi tùy theo
hoạt động của sinh vật và khác
biệt từ một phần của hệ cơ
quan nầy đến hệ khác.
Ðể tiện lợi, người ta thường
chia những chất hòa tan nầy thành sáu
loại: (1) các ion vô cơ và muối; (2) các
protein huyết tương; (3) các chất dinh dưỡng
hữu cơ; (4) các sản phẩm thải có
nitơ; (5) các sản phẩm đặc biệt
được chuyên chở; (6) các khí hòa tan.
Nồng độ của từng ion trong
huyết tương được duy trì
hằng định và được điều
hòa nhờ nhiều yếu tố, đặc
biệt là thận và các cơ quan bài tiết khác
cũng như một số hormone.
Sự ổn định nầy được
gọi là sự cân bằng nội môi
(homeostasis), đặc biệt cần thiết cho
các chức năng của cơ thể. Khi
nồng độ của các ion trong huyết tương
tăng sẽ dẫn đến sự tăng các
ion nầy trong dịch mô, gây ra những rối
loạn nghiêm trọng.
Nồng độ của các ion nầy cũng
rất quan trọng trong việc xác định
độ pH của dịch cơ thể. (2)
Các protein huyết tương chiếm khoảng
7 - 9% trọng lượng huyết tương,
gồm ba loại chính: fibrinogen, albumin và globulin,
hầu hết đều được tổng
hợp từ gan. Các
protein nầy có vai trò quan trọng trong việc xác
định áp suất thẩm thấu của
huyết tương, ảnh hưởng đến
sự trao đổi chất ở mao mạch và
sự cân bằng nước của cơ
thể. Chúng giúp
ổn định pH của huyết tương
cũng như kiểm soát độ nhớt
của huyết tương.
Hình
1. Nguồn
gốc của các tế bào máu
Ngoài ra, khi liên kết với các hormone, acid
béo hoặc các lipid, một số vitamin và các
chất khoáng, các protein sẽ hổ trợ cho
sự vận chuyển các chất nầy
bởi máu. Thêm
vào đó, fibrinogen và một số globulin có vai
trò trong sự đông máu, một số globulin
khác tham gia vào đáp ứng miễn nhiễm.
(3) Các chất hữu cơ trong huyết tương
gồm glucoz, các chất béo, phospholipid, acid amin,
acid lactic và cholesterol.
Một số được hấp thu
từ ruột, một số đi vào máu từ
gan. Acid lactic là
sản phẩm của sự đường phân,
chúng được chuyên chở từ máu vào
gan. Tại đây
một số được dùng để tái
tổng hợp carbohydrate, một số sau đó
được oxy hoá thành
(4) Huyết tương cũng chuyên chở
các sản phẩm thải có nitơ từ các cơ
quan bài tiết như thận.
Ở động vật hữu nhũ,
những chất thải nầy chủ yếu
ở dạng ure, một số ít là ammonia và
acid uric.
(5) Trong số các sản phẩm được
huyết tương chuyên chở, các hormone có
vai trò đặc biệt quan trọng.
Cấu trúc, chức năng và cơ chế
tác động của chúng đã được
đề cập chi tiết ở chương 7. (6)
Có ba chất khí chính hòa tan trong huyết tương.
Một là N2 khuếch tán từ phổi vào
máu, trơ về mặt sinh lý.
Hai khí khác là
b. Bạch cầu Các
tế bào bạch cầu của người có
nhân lớn, hình dạng không đều (Hình1).
Chúng được tạo ra từ các nguyên
bào đặc biệt trong tủy xương và
được phóng thích vào dòng máu.
Ngoài máu, bạch cầu còn có rất
nhiều trong hệ bạch huyết.
Chúng cũng có khả năng di chuyển
tự do trong các mô liên kết.
Một số có chuyển động
kiểu amip và có thể thoát ra khỏi mạch
máu và mạch bạch huyết bằng các xuyên
qua thành mạch ở chỗ tiếp giáp
giữa các tế bào nội bì.
Thực chất các tế bào bạch
cầu di chuyển trong một hệ thống liên
tục bao gồm máu, bạch huyết và các mô
liên kết. Các
tế bào bạch cầu khác nhau giữ vai trò
rất quan trọng trong việc bảo vệ cơ
thể chống lại các vi sinh vật gây
bệnh như chúng ta sẽ thấy trong phần
sau. c.
Hồng cầu Các
hồng cầu của người là những
tế bào nhỏ, hình đĩa lõm hai mặt,
không có nhân. Ở
cá thể trưởng thành, các hồng cầu
được sản sinh từ các nguyên bào
trong tủy xương (Hình 1).
Các tế bào hồng cầu chưa trưởng
thành có nhân, ti thể, bộ Golgi...
nhưng về cuối giai đoạn phát
triển, chúng mất nhân và các bào quan khác, tích
tụ nhiều hemoglobin, sau đó đi vào máu.
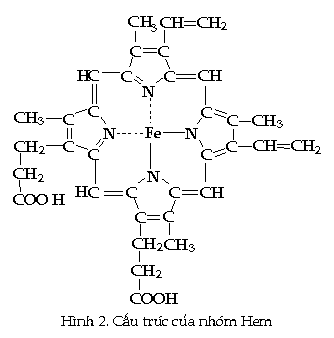
Phân tử Hemoglobin (Hb) là một protein hình
cầu có bốn chuỗi polypeptide. Mỗi
chuỗi đều có chứa một nhóm
phụ phức tạp gọi là nhóm Hem, có
một nguyên tử Fe ở trung tâm (Hình 2). Mỗi
nguyên tử Fe
của Hb có
thể kết hợp với một nguyên tử
oxy. Ở
phổi, nồng độ oxy trong môi trường
chung quanh tương đối cao, mỗi phân
tử Hb kết
hợp với bốn
phân tử oxy tạo thành
một hợp chất
là oxyhemoglobin
làm cho máu có màu đỏ tươi. Trong mao
mạch của hệ tuần hoàn, nơi có
nồng độ oxy tương đối
thấp, Hb sẽ phóng thích oxy. Khả năng máu
chuyên chở
d.
Tiểu cầu Tiểu
cầu là những thể nhỏ, không màu, có
nhiều hạt, kích thước nhỏ hơn
hồng cầu rất nhiều. Tiểu cầu
được sản sinh ra khi tế bào
chất của các tế bào tủy xương
(megakaryocyte) bị tách ra và đi vào hệ
tuần hoàn.
Chức năng chính của tế bào là
giải phóng Thromboplastin để gây đông máu.
Khi gặp một vật lạ hay bề
mặt tiếp xúc nhám, tiểu cầu sẽ ngưng
kết thành cục nhờ đó đóng kín
vết thương. Ngoài
ra khi tiểu cầu bị vỡ chúng sẽ phóng
thích serotonin gây co mạch để cầm máu. Sự
đông máu là một sự thích nghi tiến hóa
cho sự sửa chữa cấp thời của
hệ tuần hoàn và để ngăn cản
sự mất quá độ của dịch cơ
thể khi mạch máu bị tổn thương.
Sự đáp ứng tức thời của
mạch máu là khép lại, làm cho máu chảy
chậm lại. Các
tiểu cầu ở vùng nầy cũng dính vào
nhau và dính vào mô tổn thương, tạo ra
một đám tiểu cầu bị ngưng
kết. Ðám
tiểu cầu có thể làm chậm hoặc làm
ngừng chảy máu từ các mạch tổn thương
nhưng chúng rất dễ bị đẩy ra
khỏi vị trí. Chúng
được ổn định bằng sự
thành lập của một cục máu (một
mạng lưới các sợi được
tạo ra chung quanh các tiểu cầu ở các mô
tổn thương).
Các tế bào khác có thể đan xen vào các
sợi làm căng cục máu.
Các sợi nầy được hợp thành
từ các protein fibrin.
Sợi fibrin được thành lập
trong quá trình đông máu khi một protein tan trong
huyết tương là fibrinogen được
biến đổi thành fibrin không hòa tan.
Mặc dù quá trình nầy rất phức
tạp và gồm hàng loạt phản ứng,
để đơn giản nó có thể
được tóm gọn lại trong hai phản
ứng sau:
Quá
trình bắt đầu khi bề mặt của
mạch máu bị tổn thương phóng thích
ra một chất gọi là thromboplastin, chất
nầy kết hợp với các protein khác
của máu tạo thành một phức hợp
được hoạt hóa.
Phức hợp nầy biến đổi
protein của huyết tương là prothrombin thành
thrombin. Ion
Nếu chẳng may một trong các yếu
tố cần thiết cho quá trình đông máu
bị thiếu, toàn bộ chuỗi phản
ứng có thể bị hỏng.
Thí dụ những người mắc
bệnh máu khó đông (hemophilia) thiếu yếu
tố VIII là
một loại protein cần cho sự thành
lập phức hệ hoạt động để
biến đổi prothrombin thành thrombin.
Không có protein nầy máu không đông
được, do đó ngay cả một
vết đứt nhỏ cũng làm cho người
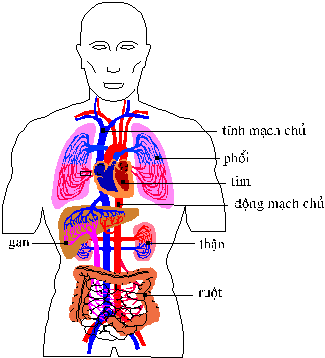
bệnh chảy máu đến chết. Người cũng như các động vật hữu nhũ có một hệ tuần hòa kín, bao gồm một tim, một số động mạch, mao mạch và tĩnh mạch . Các động mạch (artery) là những mạch chuyên chở máu từ tim đi ra, trong khi tĩnh mạch (vein) mang máu trở về tim. Cần lưu ý rằng cơ sở để phân biệt động mạch và tĩnh mạch là chiều di chuyển của máu chứ không phải là hàm lượng oxy trong máu. Các mao mạch (capillary) là những mạch máu rất nhỏ nối liền giữa động mạch và tĩnh mạch . Chúng thường chạy từ các tiểu động mạch (arteriole) đến các tiểu tĩnh mạch (venule). Các mao mạch rất quan trọng vì phần lớn sự trao đổi chất giữa máu và các mô khác diễn ra qua lớp thành mỏng của chúng. Bây
giờ chúng ta hãy theo dõi sự di chuyển
của máu trong hệ tuần hoàn của người
bắt đầu với máu trở về tim
từ tay hoặc chân.
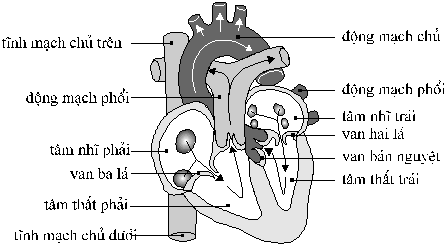
Máu này đi vào buồng trên, bên phải
của tim, gọi là tâm nhĩ phải (right
atrium) theo một trong hai tĩnh mạch
lớn là tĩnh mạch
chủ trên hoặc tĩnh mạch chủ dưới
(anterior or posterior vena cava).
Sau đó máu di chuyển qua một van
gọi là van ba lá (tricuspid valve) đi vào tâm
thất phải (right ventricle) là buồng bên
phải ở phần dưới tim.
Ở đây do máu này trở về tim
từ mô, có ít
Hình
3. Tim người Sau
khi xuất phát từ phần trước
của tim, động mạch
chủ tạo thành một cung chạy vòng
ra phía sau theo thành sống lưng giữa
ngực và bụng. Dọc
theo chiều dài động mạch
chủ phát sinh nhiều nhánh động
mạch mang máu
đi khắp nơi của cơ thể.
Mỗi động mạch
lại phân nhánh thành các động
mạch nhỏ
hơn cho đến khi các động mạch
nhỏ nhất nối với các mao mạch trong
mô. Tại đây
Hình
4. Mô hình hệ tuần hoàn của người Thành
của động mạch và tĩnh mạch
được cấu tạo từ ba
lớp: (1) lớp ngoài cùng là mô liên kết có
nhiều sợi làm cho động mạch
có tính đàn hồi; (2) lớp giữa là
cơ trơn có thể làm thay đổi kích thước
mạch máu; (3) lớp trong cùng là mô liên kết
cùng với nội bì.
Hai lớp mô liên kết phía ngoài và phía
trong của thành động mạch
và tĩnh mạch
kết thúc ở đầu tận cùng làm
cho mao mạch chỉ có một lớp tế bào
nội bì mỏng (Hình 5).
Thành
của động mạch
và tĩnh mạch
không thấm đối với các chất
trong máu và dịch mô nên hầu như không có
sự trao đổi chất ở động
mạch hoặc
tĩnh mạch. Ngược
lại, qua lớp thành mỏng của mao
mạch sự trao đổi chất diễn ra.
Hình
5. A) Ảnh hiển vi điện tử một
động mạch (trái) và một tĩnh
mạch (phải) B)
Sơ đồ các lớp của thành động
mạch, tĩnh mạch và mao mạch
a.
Hoạt động của tim (
Chu kỳ tim: Mặc dù tim người có sự
phân chia hoàn toàn thành hai nửa trái
và phải nhưng cả hai đều đập
giống nhau theo một trình tự nhất định.
Nhờ thứ tự co bóp lần lượt
của các buồng tim và sự có mặt
của các van tim, chỉ cho máu dồn theo
một chiều, nên máu mới chảy được
thành vòng. Mỗi chu kỳ tim bắt đầu
bằng pha co đồng thời của hai tâm nhĩ
trái và phải. Máu
không dồn trở lại tĩnh mạch mà trào
xuống hai tâm thất vì sức ép lúc co tâm nhĩ
đã làm tắt lỗ thông với tĩnh
mạch, đồng thời mở các van ba lá và
van hai lá (Hình 6A và B). Ngay sau đó, hai tâm
thất cùng co, máu không dồn trở lại hai
tâm nhĩ mà tràn vào động mạch chủ
và động mạch phổi, vì sức ép lúc
co tâm thất đã làm đóng các van nhĩ
thất đồng thời làm mở các van bán
nguyệt (Hình 6C và D).
Hình
6. Tóm tắt các
sự kiện của một chu kỳ tim *Tính
tự động của tim Các
tế bào cơ tim có tính tự động nghĩa
là chúng có khả năng tự đập theo
một kiểu riêng, không cần sự kích thích
từ hệ thần kinh.
Nếu tất cả các liên hệ thần
kinh đến tim bị cắt bỏ, tim vẫn
tiếp tục đập bình thường,
mặc dù nhịp đập có thể thay đổi.
Tuy nhiên nhịp
đập của
tim cũng được điều hòa một
phần nhờ các kích thích từ
hai dây thần kinh, một làm tăng và
một làm giảm nhịp đập của tim.
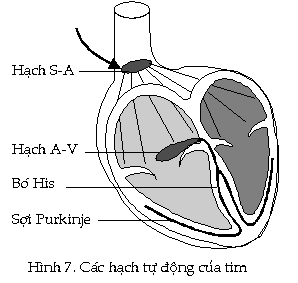
Sự khởi đầu của nhịp
đập thông thường đến từ
hạch xoang-nhĩ còn gọi là hạch S-A
(sino-atrial node). Nó
là Ở
mức điều hòa bên trong, hạch S-A
tạo ra một xung hưng phấn lan truyền
qua thành của hai tâm nhĩ làm tâm nhĩ co
lại. Khi xung
nầy truyền đến hạch A-V, hạch
sẽ bị kích thích và xung hưng phấn
sẽ nhanh chóng lan truyền từ nó đến
tất cả các phần của tâm thất theo
các sợi của bó His.
Những xung nầy sẽ kích thích tâm
thất co bóp. *
Tiếng tim và điện tim Nhịp
đập của tim (số lần tim co bóp trong
một phút) ở người bình thường
trong trạng thái nghỉ là
70 lần /phút nhưng
thay đổi
tùy theo từng cá thể.
Trong lúc đập, tim phát ra những âm
thanh đặc biệt, có thể nghe được
dễ dàng bằng một ống nghe (stethoscope).
Ðầu tiên là một âm dài, trầm
được tạo ra do sự đóng của
các van nhĩ thất khi tâm nhĩ co.
Tiếp theo là một âm ngắn, đục
và cao được tạo ra do sự đóng các
van bán nguyệt. Tiếng
tim bình thường là một dấu hiệu báo
cho các bác sĩ biết rằng tất cả các
van đều hoạt động thích hợp.
Nếu một van bị tổn thương
và không thể đóng lại hoàn toàn, một
âm thanh như tiếng gió thổi có thể nghe
được khi máu rò rĩ ngược qua van
nầy. Van tim càng
tổn thương nhiều thì hoạt động
của tim càng kém hiệu quả.
Khi tim co, cơ tim trải qua một loạt
thay đổi điện thế.
Những thay đổi nầy có thể
được phát hiện bằng cách cắm
điện cực vào da và ghi lại bằng
một dụng cụ gọi là điện tâm
kế (electrocardiograph).
Những bất thường trong hoạt
động của tim sẽ làm thay đổi
dạng của điện tâm đồ
(electrocardiogram).
Ở trạng thái nghỉ ngơi, tim
của người trưởng thành bơm
khoảng 5 lit máu/phút, tương đươmg
với tổng lượng máu trong cơ
thể. Máu lưu
thông trên một đoạn ngắn như máu
cung cấp cho cổ, ngực có thể trở
về tim nhanh và tuần hoàn nhiều lần
trong một đoạn ngắn như máu cung
cấp cho cổ, ngực có thể trở
về tim nhanh và tuần hoàn nhiều lần
trong 1 phút, trong khi máu đến các phần xa hơn
của cơ thể như chân có thể mất
nhiều phút để trở về tim.
Sự kết hợp giữa nhịp đập
của tim và sự tăng lượng máu
mỗi lần đập có thể làm tăng
tổng lượng máu được bơm
trong một phút lên gấp 4 - 7 lần so với
lúc nghỉ. b.
Huyết áp và tốc độ của dòng máu *
Huyết áp Khi
tâm thất trái co bóp, nó tống máu với áp
suất cao vào động mạch
chủ và máu dồn về hướng các
động mạch .
Thành của động mạch
đàn hồi và sóng co bóp làm căng chúng.
Lúc tim giãn, tim không tạo ra áp lực trên
máu động mạch
nên áp suất trong động mạch
giảm nhưng các sợi đàn hồi
của thành động mạch
vẫn duy trì một ít áp suất.
Như vậy có một chu kỳ đều
đặn của áp suất trong các động
mạch lớn,
đạt cao nhất lúc tim co và thấp
nhất lúc tim giãn.
Ở người, huyết áp động
mạch trong
tuần hoàn hệ thống thường
được đo ở phần trên cánh tay.
Vào giai đoạn nghỉ ở người
nam trưởng thành huyết áp tối đa (lúc
tim co) là 120 mmHg và huyết áp tối thiểu (lúc
tim giãn) là 80 mmHg. Huyết
áp giảm liên tục khi máu di chuyển càng lúc
càng xa tim. Huyết
áp cao nhất
ở động mạch
chủ trong phần nối với tim,
giảm đều ở những phần xa hơn
của động mạch
chủ và các nhánh động mạch,
giảm rất nhanh ở các động mạch
nhỏ và mao mạch.
Nó tiếp tục giảm (mặc dù
chậm hơn) trong tĩnh mạch
và thấp nhất ở các tĩnh mạch
chủ gần tim.
(Hình 8). Sự
giảm huyết áp trong các phần kế
tiếp của hệ tuần hoàn là kết
quả của sự ma sát giữa dòng máu và thành
mạch. Một
khuynh độ như thế của áp suất là
cần thiết nếu máu chảy liên tục vì
chất dịch chỉ có thể di chuyển
từ vùng có áp suất cao đến vùng có áp
suất thấp hơn.
Hình
8. Biểu đồ
áp suất máu ở những phần khác nhau trong
hệ tuần hoàn của người *
Tốc độ dòng máu Dọc
theo lộ trình của dòng máu có nhiều thay
đổi xảy ra.
Thứ nhất, do sự gia tăng
khoảng cách từ tim, khoảng cách giữa áp
suất tim co và áp suất tim giãn giảm
xuống vì độ đàn hồi của thành
động mạch có khuynh hướng làm
giảm biên độ của huyết áp.
Thứ hai, tốc độ của dòng máu
có khuynh hướng giảm khi máu di chuyển
qua các nhánh động mạch
và các động mạch
nhỏ, tốc độ thấp nhất
ở mao mạch và lại tăng lên trong các tĩnh
mạch nhỏ
và tĩnh mạch . Những
thay đổi nầy trong tốc độ dòng
máu là kết quả của sự thay đổi
trong tổng bề mặt cắt ngang của
hệ thống mạch.
Tốc độ của dòng máu tỉ
lệ nghịch với bế mặt cắt
ngang: bề mặt cắt ngang càng lớn,
tốc độ dòng máu càng chậm.
Khi các động mạch
phân nhánh thành các động mạch
nhỏ rồi thành các mao mạch, tổng
bề mặt cắt ngang tăng lên và tốc
độ dòng máu chậm lại.
Khi các mao mạch hợp thành các tĩnh
mạch nhỏ
rồi thành các tĩnh mạch, tổng bề
mặt cắt ngang lại giảm xuống và
tốc độ dòng máu tăng lên. Vì
huyết áp trung bình trong tĩnh mạch rất
thấp, khoảng 2 mmHg ở tĩnh mạch
cửa gần tim nên phải có cơ chế khác
hơn là huyết áp để đưa máu
trở về tim, đặc biệt là máu ở
những phần cự thấp phải di
chuyển ngược chiều trọng lực.
Các cử động của cơ hoành và
lồng ngực trong quá trình hô hấp góp
phần lớn vào việc đưa máu tĩnh
mạch về
tim. Khi hít vào, cơ
hoành co đồng thời lồng ngực
mở rộng ép lên xoang bụng.
Aïp suất âm được tạo ra trong
lồng ngực khi hít vào làm cho các tĩnh
mạch ở
ngực mở rộng, kéo máu vào và đẩy
máu về tim. Ðồng
thời sự nén của xoang bụng làm tăng
áp suất ở vùng đó, đẩy máu lên tĩnh
mạch cửa
sau. Thành tĩnh
mạch tương
đối mỏng và dễ đàn hồi.
Lúc cơ thể cử động, các cơ
nằm cạnh tĩnh mạch
co lại ép vào tĩnh mạch , đẩy
máu trong tĩnh mạch
đi lên. Máu
chỉ có thể di chuyển về tim vì trong tĩnh
mạch có các
van một chiều ngăn không cho máu chảy ngược
lại. c.
Chức năng của mao mạch Các
mao mạch xâm nhập vào tất cả các
phần của mọi mô. Người ta ước
lượng rằng mô cơ có khoảng 60000 mao
mạch/1cm2 của một lát cắt ngang. Ðường
kính của mao mạch rất nhỏ, xấp
xỉ đường kính của những
hồng cầu di chuyển ngang qua chúng. Sự phân
nhánh rộng rãi và đường kính nhỏ
của mỗi mao mạch không chỉ cho phép chúng
đi đến tất cả các phần
của toàn bộ các mô mà còn đảm
bảo một bề mặt thích hợp có
lợi cho quá trình trao đổi. Sự phân nhánh
cũng làm tăng tổng bề mặt cắt
ngang của hệ thống làm cho dòng máu di
chuyển trong mao mạch chậm hơn nhiều
so với trong động mạch hoặc
tỉnh mạch. Sự di chuyển chậm
của dòng máu cho phép quá
trình trao
đổi xảy
ra nhiều lần. Lòng mao mạch rất nhỏ
cũng giúp cho thời gian có lợi trong sự
trao đổi, nhờ sự tăng độ ma
sát của dòng máu và sự giảm huyết áp
trong buồng mao mạch. Sự
trao đổi vật chất giữa máu trong mao
mạch và dịch mô ngoài mao mạch xảy ra
theo ít nhất là một trong ba cách: (1) vật
chất có thể di chuyển hoàn toàn bằng
sự khuếch tán qua màng của một tế
bào nội bì trên thành mao mạch, ngang qua tế
bào chất của tế bào, và đi ra ngoài qua
màng tế bào ở phía đối diện; (2)
sự vận chuyển có thể xảy ra thông
qua một số lượng lớn các không bào
trong các tế bào nội bì. Các không bào nầy
thu nhận vật chất bằng sự nội
nhập bào ở một phía của tế bào,
di chuyển ngang qua tế bào và sau đó phóng thích
vật chất bằng sự ngoại xuất bào
ở phía đối diện; (3) khoảng
trống giữa các tế bào kế cận nhau
(khe = cleft) trong mao mạch của hầu hết các
phần cơ thể cho phép sự thấm
lọc nước và phần lớn các chất
hòa tan, trừ protein. (Hình 9). Cần lưu ý
rằng cơ chế vận chuyển trong không bào
và sự thấm lọc giữa các tế bào không
đòi hỏi sự vận chuyển của các
chất qua màng tế bào.
Hình 9. Ảnh hiển vi điện tử lát
cắt ngang qua mao mạch
Trái: Hai tế bào nội bì
tạo nên thành mao mạch. Cần lưu ý
các túi ẩm bào (mũi tên) có thể chuyên
chở các chất từ trong mao mạch ra ngoài
và ngược lại.
Phải: Ảnh phóng to thành của một
mao mạch cho thấy khe giữa hai tế bào
nội bì (mũi tên) cũng như các túi
ẩm bào. Chúng
ta hãy khảo sát quá trình trao đổi chi
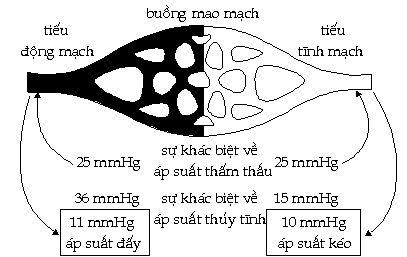
tiết hơn. Ở
cuối động mạch nhỏ của
một buồng mao mạch, tính trung bình thì áp
suất của máu cao hơn áp suất của
dịch mô ngoài mao mạch khoảng 36mmHg.
Áp suất được tạo ra bởi
các chất dịch như máu
và dịch mô, tác động vào thành của mao
mạch được gọi là áp suất
thủy tĩnh (hydrostatic pressure).
Áp
suất thủy tĩnh của máu do hoạt
động bơm của tim tạo ra.
Do độ kháng ma sát của mao
mạch và sự gia tăng bề mặt cắt
ngang, áp suất này giảm xuống còn 15mmHg lúc
máu đi đến tận cùng tĩnh mạch
của buồng mao mạch.
Áp suất thủy tĩnh của máu có xu hướng
đẩy các chất ra khỏi mao mạch vào
dịch mô chung quanh. Nếu
chỉ có một lực nầy, sẽ có
một sự mất đều đặn nước
và những chất hòa tan từ máu do sự
thấm lọc qua các khe của thành mao mạch.
Tuy nhiên, có tương đối ít sự
mất nước từ máu trong mao mạch.
Rõ ràng là có một vài lực tác động
ngược với lực thủy tĩnh.
Lực nầy xuất phát từ sự khác
biệt giữa nồng độ thẩm
thấu của máu và dịch mô.
Máu của động vật hữu nhũ
có nồng độ protein tương đối
cao (7 - 9 g/100ml) và những đại phân tử
nầy không thể dễ dàng đi qua được
thành mao mạch. Trong
dịch mô có cùng loại protein nhưng nồng
độ thấp hơn nhiều (2 g/100ml).
Do sự chênh lệch nồng độ
protein ở hai phía của thành mao mạch, máu và
dịch mô có áp suất thẩm thấu khác
nhau. Bình thường
áp suất thẩm thấu của máu cao hơn
của dịch mô khoảng 25 mmHg. Vì
vậy máu là ưu trương so với
dịch mô, làm cho nước có xu hướng
đi từ dịch mô vào máu bằng sự
thẩm thấu.
Như vậy trong khi áp suất thủy tĩnh
của máu (tạo ra bởi tim) có xu hướng
đẩy nước ra khỏi mao mạch thì
áp suất thẩm thấu lại có khuynh hướng
kéo nước vào mao mạch.
Sự di chuyển của nước sẽ
được xác định bằng dộ
lớn tương đối của hai lực
đối kháng nầy.
Cần lưu ý rằng tại tận cùng
động mạch của buồng mao mạch,
sự chênh lệch áp suất thủy tĩnh là
36 mmHg và sự chênh lệch áp suất thẩm
thấu là 25 mmHg. Làm
bài toán trừ chúng ta thấy rằng có một
áp suất khoảng 11 mmHg có khuynh hướng
đẩy nước ra khỏi mao mạch.
Ở tận cùng tĩnh mạch của
buồng mao mạch, sự chênh lệch áp
suất thủy tĩnh giảm xuống còn 15
mmHg, trong khi sự chênh lệch áp suất
thẩm thấu hầu như không đổi (dĩ
nhiên, sự mất nước của máu làm tăng
nhẹ nồng độ của protein trong máu và
kéo theo sự gia tăng áp suất thẩm
thấu nhưng sự thay đổi nầy tương
đối ít và có thể bỏ qua).
Vì vậy có một áp suất khoảng 10
mmHg có xu hướng kéo nước vào trong mao
mạch (Hình 10).
Hình
10. Sơ đồ
trình bày cơ chế của sự lọc các
chất qua thành mao mạch
Tóm
lại sự cân bằng giữa áp suất
thủy tĩnh của máu và áp suất thẩm
thấu là yếu tố để nước
được đẩy ra khỏi mao mạch
tại các tận cùng của động
mạch và được kéo vào mao mạch
tại các tận cùng của tĩnh mạch.
Vì nước có mang theo các phân tử
của nhiều chất hòa tan, chúng ta có thể
nói rằng máu trong mao mạch trước tiên
nhả các chất cho mô tại tận cùng động
mạch và sau đó thu nhận các chất để
vận chuyển từ tận cùng tĩnh
mạch. Trong quá
trình nầy, thường chỉ có một ít nước
bị mất đi từ máu.
húng
ta đã thấy rằng phần lớn các
chất dịch rời khỏi mao mạch
tại tận cùng của các động
mạch nhỏ và thường được tái
hấp thu ở tận cùng của các tĩnh
mạch nhỏ. Nhưng điều gì xảy ra
đối với phần dịch còn lại? Có
thể chúng được đưa trở
lại máu bằng những phương tiện
khác hơn sự tái hấp thu trực tiếp vào
các mao mạch máu? Ðúng vậy, các động
vật có xương sống có một hệ
đặc biệt gọi là hệ bạch
huyết giữ nhiệm vụ đưa nước
và các chất hòa tan từ mô về máu. Hệ
bạch huyết gồm một mạng lưới
các mạch được phân bố rộng rãi
khắp các phần của cơ thể.
Những mạch nầy bao gồm các tĩnh
mạch và các mao mạch bạch huyết. Các
mao mạch bạch huyết là những mạch
rất nhỏ, bịt đầu, nằm ở các
khoảng gian bào. Dịch mô có protein và các
chất khác được hấp thu vào mao
mạch bạch huyết. Các mao mạch nầy
tập trung lại thành các tĩnh mạch
bạch huyết nhỏ, sau đó tiếp
tục hợp nhất thành các tĩnh mạch
bạch huyết lớn hơn và cuối cùng là
hai ống bạch huyết rất lớn đổ
vào tĩnh mạch lớn
của hệ tuần hoàn máu ở phần trên
của ngực, gần tim (Hình 11).
Các mao mạch bạch huyết có tính
thấm cao đối với protein; tất
cả các protein bị rò rĩ khỏi mao
mạch máu có thể khuếch tán vào các mao
mạch bạch huyết để được
đưa về máu. Quá trình nầy rất quan
trọng trong việc duy trì cân bằng thẩm
thấu bình thường giữa máu và dịch
mô.
Bên cạnh việc đưa dịch mô và
protein thừa về máu, hệ bạch huyết
còn tiến hành nhiều chức năng khác. Nó
giữ vai trò chính trong đáp ứng miễn
nhiễm như chúng ta sẽ thấy ở
phần sau của chương. Các mạch
bạch huyết cũng thu nhận các chất béo
được hấp thu từ ruột.
Ngoài các mạch bạch huyết, ở
động vật hữu nhũ còn có các
hạch bạch huyết. Chúng nằm dọc theo
các mạch bạch huyết chính và được
tạo thành từ một mạng lưới các
mô liên kết. Hạch là nơi trú ẩn
của nhiều tế bào bạch cầu
thực bào. Khi bạch huyết di chuyển qua
hạch, nó được lọc và những
phần tử như các tế bào chết, các mãnh
vở tế bào, các tế bào ung thư và các
vi khuẩn bị nhốt lại và bị phá
hủy bởi các tế bào thực bào. Các
phần tử như bụi không bị các
tế bào thực bào phá hủy sẽ được
trử lại trong hạch. Vì những hạch
nầy hoạt động trong suốt quá trình
viêm nhiễm, chúng thường bị sưng lên
và gây đau nhức như trường hợp
các hạch hạnh nhân sưng lên khi cổ
họng bị viêm. Vì
hệ bạch huyết không nối với động
mạch của hệ tuần hòan máu, bạch
huyết không thể di chuyển do áp lực
xuất phát từ tim. Chuyển động
của bạch huyết giống như chuyển
động của máu trong tĩnh mạch, là
kết quả của sự thay đổi áp
suất được kích thích bởi cử
động hô hấp và do sự co của các cơ
vân ép lên thành của mạch bạch huyết
đẩy bạch huyết về phía trước
qua các van một chiều. Cơ
thể động vật và người thường
bị bao vây bởi vô số mầm bệnh. Các
vi khuẩn, virus, nấm và các sinh vật ký sinh
có mặt khắp nơi: trong thức ăn, nước
uống, không khí, đất, thậm chí trong cơ
thể sinh vật. Các vi sinh vật nầy có
thể gây nguy hiểm khi chúng xâm nhập vào cơ
thể. May thay, ở người có một
hệ thống bảo vệ (hệ miểm
nhiễm) hoạt động phá hủy các vi
sinh vật từ ngoài xâm nhập. Ở người
hệ miễn nhiễm bao gồm nhiều cơ
quan như tuyến ức (thymus), các hạch
bạch huyết, tủy xương, tụy
tạng và ruột thừa. Tất cả các
tế bào miễn nhiễm xuất phát từ các
nguyên bào trong phôi, sau đó di chuyển đến
các mô và cơ quan chuyên
biệt, tại đây chúng sản sinh ra các
hồng cầu và bạch cầu.
Hồng cầu mang oxy cho máu và
không cóï
vai trò miễn nhiễm. Những tế bào
bạch cầu nhất là các đại thực
bào, tế bào B, T, NK và tế bào mast giữ vai
trò quan trọng nhất trong đáp ứng
miễn nhiễm.
Hầu như tất cả động
vật đều cóï
các tế bào thực bào để tiêu hóa vi
khuẩn và các tế bào chết.
Những đại thực bào này bị lôi
cuốn bởi các hóa chất phóng thích từ các
mô bị bịnh hoặc các tế bào lạ;
tại đó chúng tạo ra sự viêm và
một chất nước của các tế bào
chết và các mãnh vở khác mà thường
được gọi làì mủ.
Phản ứng tự vệ tương
đối chậm và không
cóï tính chọn lọc nầy chuẩn
bị cho một hệ miễn nhiễm
cóï tính chuyên biệt
cao ở động vật
cóï xương. Cóï lẽ đó làì
sự thích nghi đối với cơ thể
cóï kích thước to và một đời
sống dài hơn. Các đáp ứng miễn
nhiễm bao gồm hai hệ thống: miễn
nhiễm qua thể dịch và miễn nhiễm
qua trung gian tế bào. Cả hai hệ thống
hoạt động chung để phá hủy các
kháng nguyên (antigen) là những chất thường
từ bên ngoài xâm nhập vào cơ thể.
Hệ thống miễn nhiễm thể
dịch bảo vệ cơ thể chống
lại các kháng nguyên thông qua sự tiết các
protein chuyên biệt gọi là kháng thể
(antibody), làm bất hoạt hoặc phá hủy kháng
nguyên trên bề mặt của vi khuẩn,
nấm, virus nằm bên ngoài tế bào, cũng như
các chất độc tự do trong máu và
huyết tương. Ngược lại, hệ
miễn nhiễm qua trung gian tế bào trực
tiếp phá hủy các vi khuẩn qua việc
sản xuất những tế bào chuyên biệt
tấn công vào các chất xâm nhập. Chúng
bảo vệ cơ thể chống lại
những mầm bệnh như virus sống trong
tế bào.
a.
Phân tử kháng thể của tế bào B Trong
quá trình phát triển phôi,
cóï hàng triệu tế bào B được
tạo ra và mỗi tế bào
cóï hàng ngàn kháng thể giống nhau
gắn trên màng tế bào.
Mỗi phân tử kháng thể có hình
chữ Y và gồm 4 sợi polypeptid, 2 sợi
"nặng" và 2 sợi "nhẹ"
nối với nhau bằng liên kết disulfide. Trình
tự amino acid của khoảng 3/4 trên mỗi
sợi nặng và khoảng 1/2 trên mỗi
sợi nhẹ là giống nhau trong tất cả
các kháng thể và tạo thành vùng không thay
đổi. Vùng còn lại của các sợi
(ở đầu tận cùng của hai nhánh
chữ Y) thay đổi rất nhiều. Vị
trí để các kháng nguyên gắn vào ( 2 vị
trí giống nhau trên mỗi phân tử kháng
thể) là ở
đầu các đoạn
cóï thể thay đổi được.
Mỗi đầu nầy là một cái túi
hay cái khe do một phần sợi nặng và
một sợi nhẹ nối với nhau.Vùng
nầy cóï
thể gắn với 6 acid amin hay các đơn
vị carbohydrate của một kháng nguyên
như kiểu enzim gắn vào các giá
thể (Hình 12).
Hình
12. Phân tử kháng thể của tế bào B b.
Sự phát triển của đáp ứng thể
dịch Hçnh 13. Sæû caím æïng lympho
baìo B do
khaïng
nguyãn
Vì
mỗi phân tử kháng thể có thể gắn
với 2 phân tử kháng nguyên, các kháng thể có
khuynh hướng làìm ngưng kết các kháng
nguyên, vi sinh vật hay siêu khuẩn mang kháng nguyên,
làm trung hòa các mầm bịnh một cách
trực tiếp.Sự ngưng kết tạo ra 3
phản ứng : (
Thứ nhất, sự ngưng kết làm cho các
đại thực bào trong bạch huyết
dễ dàng nhận diện
kháng nguyên gắn với kháng thể, và
nuốt lấy chúng.
Ðó là phản ứng với các độc
tố, siêu khuẩn, và phần lớn các vi
khuẩn. (
Thứ hai, các bạch cầu NK nhận ra các kháng
thể, bám lấìy chúng
và tiết ra các hóa chất để tiêu
diệt chúng. (
Sau cùng các kháng thể khởi động
một hệ thống bổ sung (complement system)
với hơn 20 protein huyết tương.
Trong chuỗi phản ứng nầy,
mỗi protein xúc tác làìm
hoạt hóa protein kia.
Có 4 nhóm protein tập hợp lại thành 18
kênh trên màng tế bào lạ,
nước vào tế bào theo các kênh
nầy làìm tế bào trương và vở ra (Hình
14). Hình
14. Các cách
thức kháng thể thể dịch phá hủy
mầm bệnh
a.
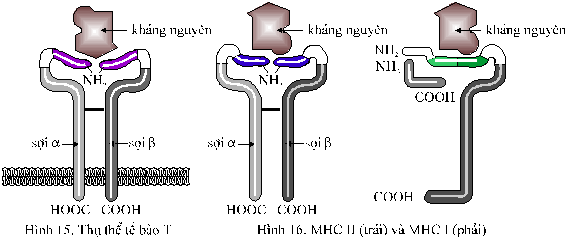
Thụ thể của tế bào T Các
tế bào T hầu hết giống như các
tế bào B nhưng chúng không có các kháng thể
gắn trên màng. Thay vào đó, mỗi tế bào
T có các thụ thể, giống như các kháng
thể nhưng chỉ có hai sợi polypeptid khác
nhau.Thụ thể
của tế bào T
không được
tiết ra, mà được gắn
chặt vào màng tế
bào bằng cái đuôi của nó. Giống
như kháng thể của tế bào B,
mỗi sợi polypeptid của
thụ thể có một đoạn
không thay đổi và một đoạn có
thể thay đổi phía ngoài cùng. Khoảng
trống giữa hai đoạn có thể thay
đổi là nơi gắn vào của các kháng
nguyên. Mỗi tế bào T
sản xuất các thụ thể
chuyên biệt
cho mỗi
một định vị kháng nguyên, và hầu
như mỗi
tế bào T có tính
chuyên biệt
độc nhất (Hình 15). Thụ
thể của tế bào T khác với các kháng
thể ở chỗ mỗi
lần chúng chỉ có thể gắn với
một kháng nguyên mà thôi. Thêm vào đó,
mỗi nhánh có
một chỗ dùng để gắn
với một vùng xác định trên màng
tế bào khác trong cơ thể. Vùng này là
một protein trên màng
do gen MHC (major histocompatibility
complex) tạo ra.
Chính phân tử MHC là
thành phần rất tích cực trong đáp
ứng tế bào T . Có 2 loại phân tử MHC:
MHC-II (tìm thấy trên màng tế bào B , tế bào
cytotoxic T và các đại thực bào) và MHC-I (tìm
gặp trong tất cả các tế bào
còn lại của cơ thể).
Phân tử MHC gồm
có 2 sợi polypeptide, mỗi sợi đều
có cả 2 đoạn thay đổi và
không thay đổi.
Phân tử MHC gắn vào kháng nguyên và
đưa nó lên bề mặt tế bào để
gắn với thụ thể
thích hợp của tế bàoT (Hình 16).
Kế đó các chỗ gắn được
củng cố
chặt chẽ bằng các glycoprotein
chuyên biệt
(như CD8 cho MHC-I, và CD4 cho MHC-II).
b.
Phát triển đáp ứng qua trung gian tế bào Khi
một tế bào bị
nhiễm một virus, vỏ protein của virus (kháng
nguyên) được gắn vào protein MHC-I trong
tế bào chất.( nếu kháng nguyên quá to
không vừa với MHC thì nó sẽ
được tiêu hóa thành mảnh nhỏ hơn).
Sau đdó phức hệ MHC-I/kháng nguyên
được chuyên chở ra bề mặt
của màng tế bào. Một
khi phức hệ MHC/kháng nguyên được thành
lập trên màng của
các tế bào bị
nhiễm, tế bào nguyên thủyT đến
gắn vào phức hệ nầy.
Ðược kích
thích tế bào T lớn lên và phân cắt để
cho ra nhiều tế bào tham gia lập tức vào
đáp ứng miễn nhiễm, đồng
thời cũng cho ra các tế bào trí nhớ làìm
cho các phản ứng kế
tiếp nhận nhanh chóng hơn. Dạng
lympho bào T được kích động là
tế bào cytotoxic T.
Nó tiêu diệt tế bào mang phức hệ
MHC-I/kháng nguyên bằng cách gắn thụ
thể vào phức hệ, chọc thủng
tế bào bị
nhiễm làìm nước
vào tế bào, trương lên và vỡ ra.
Còn các loại lympho bào T
chuyên biệt
khác - gọi là tế bào
giúp đỡ và đàn áp - phản
ứng với phức hệ MHC-II/ kháng nguyên, và
tham gia vào điều hành đáp ứng miễn
nhiễm.
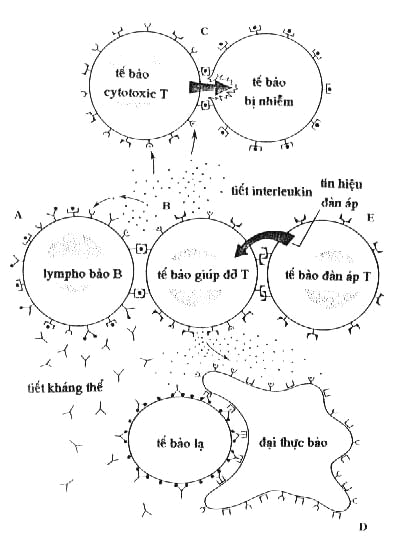
Tế bào giúp
đỡ T khởi sự điều hành đáp
ứng miễn nhiễm bằng cách gắn vào
protein MHC-II mang kháng thể đã được
xử lý thỏa đáng đến màng tế bào
của hệ miễn nhiễm. Một tế bào
giúp đỡ được hoạt hóa có
thể tiến tới khống chế kháng nguyên
bằng cách
tiết ra interleukin. Sự gắn interleukin vào các
thụ thể làìm cho số lượng tế
bào giúp đỡ
được nhân lên (Hình 17B).
Chất interleukin được
tiết ra còn
gây sự phân cắt của bất kỳ
tế bàoT xung quanh đã được hoạt
hóa bởi các kháng nguyên
chuyên biệt.
Tế bào giúp
đỡ cũng gắn vào tế bào T mang định
vị kháng nguyên thích hợp, làìm chúng tấn
công mạnh hơn các tế bào
bị nhiễm.
Khi được gắn vào lympho bào B
tế bào giúp
đỡ lại tiết
ra một loại interleukin
thứ hai làìm cho tế bào B
tiết ra kháng thể gắn vào kháng nguyên
trên tế bào gây bệnh (Hình 17A). Một
loại interleukin thứ ba cảm ứng tế bào
cytotoxic T tiêu diệt các tế bào gây bệnh (Hình
17C). Loại interleukin thứ tư cung cấp năng
lượng cho các đại thực bào xung
quanh tiêu hóa các mầm bệnh có gắn kháng
thể (Hình 17D).
Hình
17. Hoạt động
điều hành của lympho bào T
Nhiệm vụ của tế bào đàn áp
T (suppressor T cell) là tránh cho hệ miễn
nhiễm phản ứng thái quá. Tế bào đàn
áp gắn vào tế bào
giúp đỡ (theo tính
chuyên biệt
kháng nguyên) ức chế hoạt động
của tế bào nầy (Hình 17E).
Tính chuyên
biệt kháng nguyên của tế bào
giúp đỡ bắt đầu tăng lên
khi có sự gia
tăng số phức hệ MHC-II/
kháng nguyên nhưng
các tế bào đàn áp phải chờ số
tế bào giúp
đỡ tăng lên rồi
mới nhân lên và hoạt động; do
đó, đáp ứng đàn áp đi sau đáp
ứng giúp
đỡ cho tới khi tỉ lệ gặp
gỡ kháng nguyên gia tăng.
Một khi hệ miễn nhiễm bắt
đầu có
hiệu quả, hệ đàn áp đuổi
kịp và cuối
cùng khống
chế các đáp ứng. AIDS
(Acquired Immune Deficiency Syndrome)
là một bịnh tiêu hủy hầu
hết đáp ứng miễn nhiễm
bằng cách loại trừ tất cả
tế bào giúp
đỡ T quan trọng.
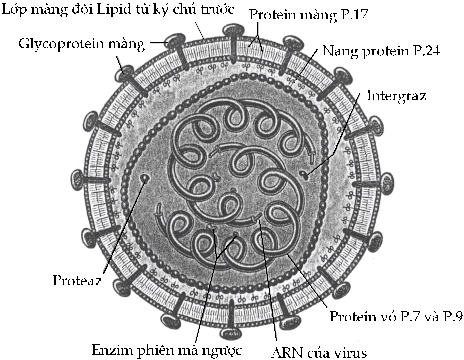
Siêu khuẩn HIV-I (human immunodeficiency virus, type I) gây
ra AIDS là loại siêu khuẩn phức tạp khác
thường. Nó
gồm có 2
sợi ARN trong bộ gen,
mỗi sợi được bao phủ
bởi một cặp protein (P7 và P9), và tất
cả được bao bọc trong hai lớp
rất khác nhau. Lớp
trong thì tương đương với vỏ
siêu khuẩn, cấu tạo bởi một
loại protein (P24). Lớp
ngoài phức tạp hơn: đó là hai lớp
lipid nhận từ một tế bào ký chủ
khi siêu khuẩn nẩy chồi,
nhưng mặt trong mang một loại
protein hướng về phía trong (P17) và một
glycoprotein (GP120) hướng ra phía ngoài.
HIV gặp
và chui vào ký chủ bằng glycoprotein GP120
gắn vào protein CD4 của tế bào
giúp đỡ T (Hình 18).
Hình
18. Cấu trúc của HIV Siêu
khuẩn HIV-1 mang 4 enzim.
Enzim 1 và 2 hoạt động chung với
nhau như enzim phiên mã ngược (reverse
transcriptase): men polymeraz tổng hợp ADN bổ
sung cho ARN sợi đơn, và men ARNpolymeraz tách
rời khuôn siêu khuẩn ra khỏi ADN bổ sung
để men polymeraz tạo ADN sợi đôi
để gắn vào bộ gen của ký chủ.
Enzim thứ ba là integraz, cắt ADN của ký
chủ và chèn các gen của siêu khuẩn vào
bộ gen của ký chủ.
Enzim thứ tư là proteaz.
Siêu khuẩn có thể gây tiêu bào
hoàn toàn hoặc bán tiêu bào (semi-lytic phage).
Trong trạng thái bán tiêu bào, siêu khuẩn sinh
sản chậm
chạp và không
giết chết ký chủ của nó.
Ðể đi đến trạng thái tiêu bào
hoàn toàn đòi hỏi phải
có cú sốc hay một sự đe dọa
với ký chủ như
có sự tham gia của nó trong đáp
ứng miễn nhiễm chẳng hạn.
Siêu khuẩn sinh sản bằng cách sao chép
trọn bộ gen của nó và cũng bằng cách
sản xuất các mARN khác nhau, một số
đa gen, để sau đó tổng hợp
trực tiếp vỏ protein GP120, loại
"P" protein, integraz và proteaz.
Protein, protein đa, và ARN tích tụ trong màng
tế bào ký chủ, chỉ
có GP120 hướng ra ngoài.
Khi các protein màng tập hợp đầy
đủ, màng bắt đầu nẩy
chồi, proteaz cắt protein đa thành phân
tử nhỏ, sản
xuất ra enzim của siêu khuẩn và protein
của vỏ. Khi
vỏ được tạo ra, siêu khuẩn
được phóng thích đi tìm một tế
bào ký chủ khác.
Ðầu
tiên, hệ miễn nhiễm phản ứng
lại siêu khuẩn, tế bào B phóng thích kháng
thể thích hợp, tế bào T nhân đáp
ứng lên, đại thực bào nuốt các siêu
khuẩn tự do. Nhưng siêu khuẩn vẫn
tiếp tục sống, ẩn nấp trong tế
bào T và không
bị tiêu hóa trong đại thực bào.
Mặc dù số lượng tế bào T có
thể duy trì bình thường khoảng một
năm nhưng các tế bào
bị nhiễm dần dần sẽ bị
tiêu bào hay bán tiêu bào.
Khi kháng nguyên GP120
xuất hiện trên bề mặt của
tế bào T, các tế bào nầy
bị tiêu diệt bởi tế bào cytotoxic
T và bạch cầu NK
nhưng việc nầy
không có ích
lợi gì cả. Không những tế bào
giúp đỡ T trong cơ thể
bị giảm đi (25% trong 3 năm sau khi
nhiễm), mà GP120 được phóng thích vào máu
và bạch huyết, nơi đây chúng gắn vào
các tế bào giúp
đỡ T khác chưa
bị nhiễm nhưng
bị tấn công và
bị giết
đi. Tệ hơn
nữa, một tế bào
bị nhiễm GP120 trên màng sẽ gắn vào
tế bào T khác và khi siêu khuẩn hòa lẫn
với ký chủ, màng tế bào
bị nhiễm sẽ hòa lẫn với
tế bào T. Hàng
tá hay hàng trăm tế bào T
bị lôi kéo vào sự hòa trộn nầy;
kết quả là tất cả đều
chết chung do bị tế bào cytotoxic
giết đi hoặc tế bào đa nhân
được tạo ra trở
nên quá to không thể hoạt động. AIDS
còn ảnh hưởng đến hệ
thần kinh. Ðại
thực bào có thể xuyên qua mạch máu vào
hệ thần kinh, nơi đây các siêu
khuẩn thoát ra, HIV nhiễm các tế bào đặc
biệt, cô
lập các neuron. Khi
các tế bào chết đi, sự dẫn
truyền trở nên
chậm chạp, kém hiệu quả, và sự chính
xác giảm đi và có thể đưa đến
dẫn truyền sai lệch. Diễn
biến ngày càng trở
nên xấu hơn.
Chẳng bao lâu số lượng tế bào
giúp đở giảm xuống 5% của
mức bình thường hay thấp hơn, và
bất cứ sự nhiễm nào cũng chỉ
gặp một đáp ứng miễn nhiễm
rất nhỏ. Bịnh
nhân AIDS thường chết trong vòng 5 đến
10 năm sau khi bị
nhiễm bịnh.
Nghiên
cứu sự lây lan của AIDS cho thấy
rằng sự lan truyền HIV hầu như luôn
luôn do máu hay chất tinh dịch của một
người nào đó vào hệ tuần hoàn
của người khác.
Do đó bịnh lan tràn do truyền máu
bị nhiễm, sử dụng lại các
kim chích, và do giao phối qua hậu môn. Siêu
khuẩn cũng có thể được
truyền qua đường âm đạo,
nhưng tỉ lệ nhiễm ở
đường nầy thấp hơn. Sự
lan truyền AIDS (hay bất cứ bịnh
truyền nhiễm nào)
tùy thuộc vào thời gian ủ trung bình
của tác nhân gây bịnh, thời gian
bị nhiễm, tỉ lệ tiếp xúc
(trong trường hợp nầy là số người
giao phối, kim chích xài đi xài lại), và
hiệu quả của mỗi lần lây
nhiễm. Trung bình từ khi
bị lây nhiễm đến khi chết
độ 8 năm. Trong
số người Mỹ đồng tính
luyến ái và người nghiện tiêm tỉnh
mạch, tỉ lệ nhiễm lên đến
70% ở các đô thị như San Francisco
và New York, là nơi có tỉ lệ tiếp xúc
rất cao; trung bình mỗi
bịnh nhân gieo bịnh cho vài người khác.
Ở những
vùng xa đô thị, sự tiếp xúc ít hơn
nên tỉ lệ thấp hơn.
Trong giao phối nam nữ, tỉ lệ
truyền bịnh và tỉ lệ tiếp xúc
thấp hơn. Hiện
nay, ở châu Âu và châu Mỹ chưa rõ
có sự truyền nhiễm trong giao phối
nam nữ (heterosexuals) hay không.
Ở châu Phi, sự lan truyền HIV
cơ bản do sự giao phối nam nữ,
và nguy cơ là do giao phối với nhiều người.
Tổ chức Y tế thế giới ước
tính ở châu Phi có gần
60% chết vì AIDS, tổng
cộng có
2,5 triệu người bị nhiễm HIV năm
1992 (và tăng lên trên
20 triệu đến cuối thế kỷ
nầy). Như vậy rõ ràng là sự giao
phối nam nữ cũng có khả năng lây lan
bịnh. Những
cố gắng đầu tiên chiến đấu
chống lại AIDS rất khó khăn là do
thiếu kiến thức về cách truyền lan
nhanh chóng của bịnh này, sự tiềm sinh dài
và chu trình phát triển của chúng.
Giờ đây thì người ta biết
được nhiều hơn, một vấn
đề rất rõ ràng là hầu như tất
cả các vaccin đều dùng kháng nguyên bề
mặt như một tác nhân giúp cho hệ
miễn nhiễm nhận biết một chứng
bịnh, phương án này lại không có tác
dụng với AIDS vì một lý do đơn
giản là nó làm cho cơ thể tiêu diệt chính
các tế bào của hệ miễn nhiễm trong
cơ thể. Việc
giấu CD4 để HIV không gắn vào cũng vô
hiệu, vì như vậy sẽ làm bất
hoạt đáp ứng miễn nhiễm.
Một trở ngại lớn nhất
với AZT là gen qui định polymeraz của HIV
có khả năng đột
biến trở nên
kháng AZT (vì gen
nầy có tỉ lệ đột biến
rất cao, 1phần 2.000 baz
bị bắt cặp sai trong bộ gen
chỉ có 10 ngàn
nucleotid). Tuy nhiên trường hợp nầy
đến nay vẫn chưa được
thấy. Thuốc
cũng rất đắt tiền và phải
sử dụng nhiều liều trong ngày.
Do đó cách chữa trị
có hiệu quả và ít tốn kém vẫn
đang được tiếp tục tìm
kiếm. Hiện
nay việc phòng ngừa bịnh bằng cách thay
đổi tập tính vẫn là tốt hơn
tất cả.
|