|
Những bài cùng tác giả Chương 19. MỐI QUAN HỆ GIỮA VIRUS VÀ TẾ BÀO Biên soạn: Phạm Văn Ty, Nguyễn Lân Dũng
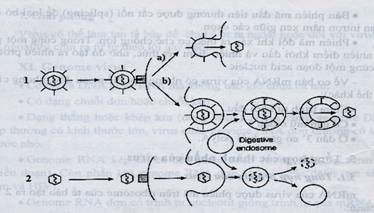
19.1. NHỮNG KHÁI NIỆM CƠ BẢN19.1.1.Mối quan hệ giữa virus và tế bàoVirus không có khả năng trao đổi chất và trao đổi năng lượng nên chúng phụ thuộc hoàn toàn vào bộ máy tổng hợp chủa tế bào. Vì vậy ở virus người ta dùng thuật ngữ nhân lên thay cho sinh sản. (1) Tế bào cho phépMuốn nhân lên virus phải xâm nhập được vào tế bào cho phép (permissive cells), đó là các tế bào có đủ điều kiện cho virus nhân lên, bao gồm: - Có thụ thể phù hợp với protein bề mặt virus. - Có các yếu tố cần thiết như yếu tố phiên mã, enzyme của tế bào. - Một số virus có phạm vi tế bào cho phép hẹp, ví dụ HBV chỉ có thể nhân lên trong tế bào gan. - Một số virus có phạm vi tế bào cho phép rộng, có thể nhân lên cả ở ĐVCXS, ĐVKXS và ở cả thực vật. - Một số chỉ nhân lên khi tế bào đang ở pha nhất định của chu kỳ phân chia tế bào. Ví dụ virus retro cần tế bào ở pha M (mitosis), khi đó màng nhân bị vỡ, genome virus mới vào được trong nhân để cài xen vào nhiễm sắc thể của tế bào. Virus parvo sử dụng enzyme DNA polymerase của tế bào chủ nên khi nhân lên chúng cần tế bào ở pha S, là lúc có mặt enzyme này. (2) Chu trình tan (lytic cycle):Chu trình nhân lên, kết thúc bằng sự làm tan và giết chết tế bào gọi là chu trình tan. Virus chỉ nhân lên theo chu trình tan gọi là virus độc. (3) Chu trình tiềm tan (lysogenic cycle):Chu trình lây nhiễm không tạo ra virus mới hay không giết chết tế bào, mà gắn xen genome của mình vào nhiễm sắc thể của tế bào, được gọi là chu trình tiềm tan. DNA của virus ở trạng thái tiềm tan gọi là provirus (nếu ở phage thì gọi là prophage), còn bản thân virus có khả năng tiến hành cả 2 quá trình tan và tiềm tan được gọi là virus ôn hoà (ví dụ phage l). Dưới tác động của yếu tố ngoại cảnh (bức xạ, hoá chất) genome virus thoát khỏi nhiễm sắc thể để tiến hành chu trình tan. 19.1.2. Tác động của virus lên tế bàoVirus có thể tác động lên tế bào theo 4 cách: (1) Gây chết tế bào. Virus nhân lên gây huỷ hoại tế bào, gọi là hiệu ứng gây huỷ hoại tế bào CPE (cytopathic effect) dẫn đến làm tan tế bào. (2) Chuyển dạng (transformation). Virus gây nhiễm nhưng không gây chết tế bào mà chuyển dạng tế nào từ trạng thái bình thường sang trạng thái biến đổi mang đặc điểm của tế bào u ác tính. (3) Nhiễm tiềm ẩn (latent infection). Virus xâm nhập và tồn tại trong tế bào ở dạng tiềm ẩn, không có tác động rõ rệt nào đến chức năng của tế bào. Những người nhiễm virus tiềm ẩn không biểu hiện triệu chứng. (4) Hấp phụ hồng cầu (haemadsorption). Một số virus có protein trên bề mặt (haemagglutinin) có khả năng kết dính hồng cầu để gây ngưng kết. Trong tế bào nuôi cấy mô, các virus này tạo ra chất ngưng kết (haemagglutinin) trên bề mặt tế bào nhiễm, nên cho hồng cầu vào chúng sẽ bị kết dính. 19.1.3. Tổng quan về quá trình nhân lên của virusQuá trình nhân lên của virus trong tế bào bao gồm 7 bước: 1- Hấp phụ (adsorption) 2- Xâm nhập và cởi vỏ (penetration và uncoating) 3- Phiên mã (transcription) tạo mRNA của virus 4- Dịch mã (translation) mRNA để tạo protein virus 5- Sao chép (replication) genome 6- Tự lắp ráp (maturation) protein với genome để tạo virion 7- Giải phóng (release) ra khỏi tế bào Nếu các bước 3, 4, 5 nhập vào một bước gọi là bước tổng hợp các thành phần (biosynthesis) thì quá trình nhân lên còn 5 bước. Ở một số virus, quá trình nhân lên không treo trình tự như trên mà xảy ra đồng thời. Ví dụ cùng lúc đều tiến hành phiên mã, dịch mã, sao chép, lắp ráp và chui ra khỏi tế bào. 19.1.3.1. Virus động vật19.1.3.1.1. Sự hấp phụVirus gắn vào thụ thể (receptor) đặc hiệu nằm trên màng sinh chất của tế bào chủ, theo nguyên tắc khoá – chìa. Một số virus còn gắn thêm vào các đồng thụ thể (co-receptor). Sự hấp phụ xảy ra tốt nhất ở 37oC. Virus gắn được vào các thụ thể là nhờ các liên kêt hoá học như liên kết hydro, ion, Vander Waals, nhưng không có liên kết đồng hoá trị. Vị trí gắn: Virus có các vị trí khác nhau chứa protein gắn vào thụ thể của tế bào. Virus trần có vị trí gắn nằm trên bề mặt capsid, đôi khi nằm sau bên trong (ví dụ virus polio) hoặc đôi khi là đỉnh của khối đa diện (virus lở mồm long móng). Ở virus adeno, vị trí gắn là đầu mút của các sợi mọc ra từ đỉnh capsid khối đa diện. Đối với virus có vỏ ngoài, vị trí gắn là các gai glycoprotein bề mặt. 19.1.3.1.2. Xâm nhập và cởi vỏSau khi gắn vào thụ thể, virus phải vượt qua màng sinh chất để xâm nhập vào tế bào theo một trong 2 cơ chế nhập bào hoặc dung hợp. 1. Nhập bào: Cả virus trần và virus có vỏ ngoài đều có thể xâm nhập theo lối nhập bào. Cơ chế chung bao gồm việc virion gắn vào màng sinh chất nơi mặt trong được bao phủ bởi lớp clathrin (protein dạng sợi tạo thành mạng lưới với các mắt hình đa giác). Virion ấn sâu vào màng tạo hốc rồi khép lại tạo thành túi nội bào hay endosome được lớp clathrin bao phía ngoài. Kênh proton của màng endosome vận chuyển H+ vào trong làm cho pH trong endosome giảm xuống ( pH 4.5 - 5). Endosome sẽ dung hợp với lysosom. Trước khi dung hợp, lớp clathrin bị loại bỏ. pH thấp hoạt hoá enzyme phân giải capsid. Cũng như màng endosome ở một số virus, pH thấp trong endosome dẫn đến việc dung hợp vỏ capsid với màng endosome, làm vỡ vỏ capsid và giải phóng genome. 2. Dung hợp: Khác với virus trần, chỉ có xâm nhập vào tế bào theo lối nhập bào, các virus có vỏ ngoài có thể vào tế bào theo cả 2 cách. Virus có vỏ ngoài có thể vào tế bào theo cách dung hợp vỏ ngoài virus với màng sinh chất vì chúng có cùng bản chất. Khi 2 màng hoà nhập sẽ đứt ra, nucleocapsid sẽ được chuyển vào tế bào chất. Sự dung hợp này xảy ra với sự tham gia của protein dung hợp (protein F). Như trên đã nói, virus có vỏ ngoài cũng có thể xâm nhập vào tế bào theo lối nhập bào. Vỏ ngoài virus bám vào thụ thể sau đó ấn lõm tạo endosome. Khi pH trong endosome giảm gai protein F sẽ chồi lên cắm vào màng endosome như chiếc neo, kéo vỏ ngoài virus sát với màng endosome và tiến hành dung hợp.
Hình 19.1: Sự nhập bào theo kiểu thực bào, tạo endosome Bơm proton làm giảm pH trong endosome, hoạt hoá sự dung hợp giữa vỏ ngoài virus với màng endosome (1b), hoặc hoạt hoá enzyme làm tan màng endosome (2). Dung hợp xảy ra trên bề mặt tế bào giữa vỏ ngoài virus với màng sinh chất. 19.1.3.1.3. Sự vận chuyển genome virus vào nhânHầu hết virus RNA ở eukaryota tiến hành sao chép trong tế bào chất, vì chúng có thể mã hoá cho tất cả các enzyme cần cho sao chép genome mà không cần đến các enzyme của tế bào nằm trong nhân. Virus cúm A là ngoại lệ, chúng cần bộ máy cắt nối của tế bào nên genome của chúng phải được đưa vào nhân. Virus retro cũng là virus RNA, nhưng tiến hành sao chép trong nhân. Thoạt đầu chúng nhờ enzyme phiên mã ngược tổng hợp DNA trên khuôn RNA trong tế bào chất, sau đó nằm đợi cho đến khi tế bào bắt đầu phân chia (giai đoạn M), màng nhân tạm thời bị vỡ, DNA cùng với protein liên kết lúc đó mới vào được trong nhân. Do đó virus này chỉ nhân lên được ở giai đoạn tế bào đang phân chia. Hầu hết virus DNA tiến hành sao chép trong nhân, ngoại trừ virus pox và irido có thể sao chép trong tế bào chất. Đối với virus sao chép trong nhân, protein cấu trúc của chúng có trình tự bám được vào vi ống (microtubule). Vi ống là các ống rỗng, đường kính 25 nm, là thành phần của bộ khung tế bào, có chức năng nâng đỡ và neo giữ nhiều thành phần của tế bào và được ví như đường ray để vận chuyển vật chất hoặc bào quan tới một vị trí nhất định trong tế bào. Một đầu vi ống được ký hiệu là (+) và một đầu ký hiệu là (-). Đầu (+) nằm gần màng sinh chất, còn đầu (-) nằm gần nhân. Một protein gọi là proetin vận chuyển (motor protein) tự nó di chuyển và chở các chất cũng như bào quan dọc theo vi ống từ vùng ngoại vi của tế bào tới vùng gần nhân. Các virus herpes, adeno, parvo, retro sử dụng hệ thống này để chở nucleocapsid vào sát màng nhân. Màng nhân được cấu tạo từ 2 lớp lipid kép, ở đó có lỗ nhân. Hầu hết nucleocapsid có kích thước quá lớn để có thể lọt qua lỗ nhân. Các phân tử muốn qua lỗ phải tạo phức với các protein chuyên biệt của tế bào có chức năng mang gọi là importin để mang vào hoặc exportin để mang ra khỏi nhân. Kênh có thể mở để cho phép các hạt có kích thước đến 25 nm thậm chí lớn hơn đi qua, ví dụ virus nhỏ như parvo, nhưng với virus lớn hơn thì phải cởi vỏ ở lỗ nhân. 19.1.3.1.4. Cởi vỏ cho genome Cởi toàn bộ hoặc một phần vỏ để giải phóng genome ra khỏi vỏ capsid. Tuỳ loại virus mà quá trình có thể diễn ra tại các vị thí khác nhau: · Trên bề mặt tế bào, capsid rỗng nằm lại bên ngoài tế bào. · Cởi vỏ bên trong tế bào chất. · Cởi vỏ tại lỗ nhân. · Cởi vỏ bên trong nhân. Cần nhớ rằng virus xâm nhập thành công vào tế bào không có nghĩa là chúng luôn luôn nhân lên được. Tế bào cũng có cơ chế bảo vệ chống lại virus, ví dụ enzyme từ lysosom có thể làm bất hoạt virus trước và sau cởi vỏ; interferon cảm ứng tạo protein độc ức chế sự nhân lên của virus. Một số virus nhiễm ở dạng tiềm ẩn, chúng không nhân lên, nhưng genome vẫn còn nguyên vẹn và vẫn có tiềm năng nhân lên. 19.1.3.1.5. Tổng hợp các thành phầnSau khi xâm nhận vào tế bào, pha đầu tiên của chu trình nhân lên gọi là pha ẩn. Ở giai đoạn này không phát hiện được bất kỳ virus nào. Đây là đặc điểm chỉ thấy có ở virus. Sự biểu hiện của genome được bắt đầu rất sớm, ngay sau khi virus xâm nhập vào tế bào. Có 4 quá trình xảy ra trước khi hạt virus được lắp ráp: (1) Phiên mã tạo mRNA (2) Dịch mã sớm tạo protein phi cấu trúc – đó là các enzyme dùng cho sao chép (3) Sao chép tạo genome (4) Dịch mã muộn tạo protein cấu trúc để cấu tạo capsid và vỏ ngoài (1) -Phiên mã tạo mRNAa- Phiên mã genome virus Thông thường phiên mã là quá trình truyền thông tin di truyền từ DNA sang RNA nhờ enzyme RNA polymerase (chính xác hơn là RNA polymerase phụ thuộc DNA), tuy nhiên genome của virus có thể là RNA, cho nên ở virus phiên mã đơn giản chỉ là quá trình truyền thông tin di truyền từ genome sang mRNA. Có 3 loại RNA polymerase: RNA polymerase I phiên mã rRNA, RNA polymerase II phiên mã các gen mã hoá protein và RNA polymerase III phiên mã rRNA 5S, tRNA và các RNA nhỏ bé khác. Dựa vào quá trình phiên mã, tạo ra mRNA mà David Baltimore đã phân loại tất cả virus thành 7 nhóm như đã nêu ở phần phân loại: (1) Virus DNA kép, (2) virus DNA đơn (+/-), (3) virus RNA kép, (4) Virus RNA (+), (5) virus RNA (-), (6) virus RNA phiên mã ngược và (7) virus DNA phiên mã ngược. Acid nucleic sợi đơn (DNA hoặc RNA) có trình tự nucleotide giống với trình tự nucleotide của mRNA quy ước là sợi (+), ngược lại có trình tự tương bù với mRNA được quy ước là sợi (-). Nhóm I. Virus DNA kép hầu hết tiến hành phiên mã trong nhân, sử dụng RNA polymerase phụ thuộc DNA (tức là RNA polymerase II) của tế bào. Đối với virus DNA kép thì không phân biệt sợi (+) và sợi (-) mà gọi là sợi trái (L) và sợi phải (R), bởi vì genome của hầu hết các virus này có khung đọc mở (ORF) theo cả 2 hướng. Một số virus DNA phiên mã trong tế bào chất (ví dụ virus pox), sử dụng RNA polymerase phụ thuộc DNA do virus mã hoá. Nhóm II. Virus DNA đơn (+) hoặc (-), tất cả đều phiên mã trong nhân và sử dụng RNA polymerase II của tế bào. Cả virus DNA (+) và DNA (-) khi phiên mã tạo mRNA đều phải qua giai đoạn trung gian DNA kép, dạng sao chép, viết tắt là RF (replicative form). Nhóm III. Virus RNA kép, tất cả đều phiên mã trong tế bào chất, sử dụng enzyme RNA polymerase phụ thuộc RNA do virus mã hoá để tạo mRNA. Nhóm IV. Virus RNA (+) có chức năng của mRNA trước khi phiên mã phải dich mã để tạo RNA polymerase phụ thuộc RNA của riêng mình sau đó mới phiên mã tạo mRNA. Nhóm V. Virus RNA (-) không phân đoạn phiên mã trong tế bào chất, sử dụng RNA polymerase phụ thuộc RNA do chúng mang theo. Virus RNA (-) phân đoạn (ví dụ virus cúm) tuy phiên mã trong nhân nhưng chúng sử dụng RNA polymerase thụ thuộc RNA mang theo, vì tế bào không có enzyme này. Nhóm VI. Virus retro phiên mã ngược, gồm 2 giai đoạn: Lúc đầu phiên mã genome RNA (+) thành cDNA (-) tiến hành trong tế bào chất nhờ enzyme phiên mã ngược của virus, sau đó DNA vào nhân tích hợp với nhiễm sắc thể của tế bào và tiến hành phiên mã tạo mRNA nhờ enzyme RNA polymerase phụ thuộc DNA của tế bào. Nhóm VII. Virus DNA kép phiên mã ngược, ví dụ HBV, tiến hành phiên mã trong nhân dùng enzyme RNA polymerase phụ thuộc DNA của tế bào để tạo RNA (+) tiền genome, sau đó ra khỏi nhân, dùng enzyme phiên mã ngược của virus để phiên mã RNA tiền genome thành DNA (-) rồi sau đó tạo genome DNA kép. Tất cả các virus có genome RNA đều phiên mã trong tế bào chất trừ virus cúm. Tất cả các virus có genome RNA (-) đều phải mang theo RNA polymerase phụ thuộc RNA để phiên mã.
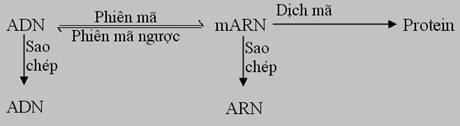
b- Cải biên lý thuyết trung tâm
Năm 1958 Francis Crick đưa ra
lý thuyết Trung tâm (Central Dogma), theo đó dòng thông tin di truyền luôn
đi từ DNA
Những kiến thức về virus đã đưa đến sự cải biến lý thuyết trung tâm. Nhiều virus có genome RNA được sao thành RNA và một số có genome RNA được phiên mã ngược thành DNA.
c- Promoter và enhancerTrong promoter của nhiều tế bào eukaryota và virus có trình tự chung (consensus) sau: TATAA/TAA/TA/G. Không phải promoter nào cũng có trình tự giống hệt nhau mà có thể sai lệch, nên gọi là trình tự chung. Trình tự TATAA còn gọi là hộp TATA hay hộp Pridnow nằm ở phía trước điểm khởi đầu phiên mã, khoảng 25-30bp. Hộp TATA chịu trách nhiệm cho phép RNA polymerase gắn vào vùng khởi động phiên mã. Ví dụ: Hộp TATA có trong promotor duy nhất của HIV-1, nhưng trong số 4 promoter của HBV thì chỉ có 1 promoter chứa hộp này. Enhancer (đoạn tăng cường) có trình tự bám vào yếu tố phiên mã. Sự tương tác này làm tăng tốc độ phiên mã bởi RNA polymerase II.
Hình 19.2: Phiên mã từ DNA kép của eukaryota d- Yếu tố phiên mãYếu tố phiên mã là các protein gắn đặc hiệu vào promoter và enhancer để kiểm soát sự biểu hiện gen. Một số virus tạo ra yếu tố phiên mã của riêng mình, ví dụ VP/6 của virus herpes simplex, nó là một thành phần của virion và protein Tax của virus HTLV (virus gây ung thư tế bào T ở người) được tạo ra trong tế bào nhiễm. Một số yếu tố phiên mã của tế bào cũng tham gia hoạt hoá hoặc kiềm chế sự phiên mã của các gen virus. Các yếu tố phiên mã đặc hiệu mô (tissue – specific transcription factors) cần cho một số virus mang tính đặc hiệu mô nghiêm ngặt, nghĩa là chúng chỉ nhân lên trong một số mô nhất định. Một số yếu tố phiên mã của tế bào có tên là yếu tố phiên mã chung (general transcription factor) tham gia vào sự kiểm soát biển hiện gen của nhiều loại tế bào và virus. Ví dụ yếu tố phiên mã TFIID bám vào hộp TATA (hình 19.3).
Hình19.3: Sự bám của các yếu tố phiên mã và RNA pol II vào hộp TATA TFIID là một phức hợp gồm 13 polypeptide, một trong số đó là protein bám hộp TATA. Sau khi TFIID bám vào hộp TATA là đến lượt các yếu tố phiên mã chung khác (TFIIA, TFIIB, TFIIE, TFÌI, TFIIH) và RNA pol II bám vào. Trong số các yếu tố phiên mã của tế bào bám vào enhancer, có: - AP1 và AP2 (activator protein 1 và 2) là các protein hoạt hoá. - SP1 (stimulator protein) là protein kích thích. - NF-kB (nuclear factor kB) là yếu tố nhân kB. Hầu hết các yếu tố phiên mã này tham gia vào quá trình phiên mã của HIV-1. Cũng như hoạt hoá sự biểu hiện gen, các yếu tố phiên mã cũng tham gia vào sự kiềm chế biểu hiện gen. Mọi cơ thể đều có khả năng tự điều hoà các gen của mình. Ví dụ côn trùng có các gen khác nhau và sự biểu hiện các gen này tuỳ thuộc việc chúng đang ở giai đoạn nào: ấu trùng, sâu non, sâu trưởng thành hay bướm. Ở virus cũng vậy, chúng có các gen khác nhau, biểu hiện ở các thời điểm khác nhau. Gen sớm mã hoá cho các protein sớm, đó là các protein không cấu trúc, thường là các enzyme và gen muộn mã hoá cho các protein muộn, đó là các protein cấu trúc, tham gia vào tạo capsid hoặc vỏ ngoài. e- Gắn mũ vào RNANgay sau khi tổng hợp RNA và trong lúc quá trình phiên mã còn đang tiếp tục thì hầu hết các bản phiên mã (RNA) đã được gắn mũ ở đầu 5’. Mũ là guanosine triphosphate nối với một nucleotide ở đầu 5’ nhờ liên kết 5’-5’, thay vì liên kết 5’-3’ như bình thường. Hầu hết mRNA ở tế bào eukaryota và ở virus đều có mũ ở đầu 5’. Mã có chức năng: · Giúp mRNA vận chuyển từ nhân ra tế bào chất. · Bảo vệ mRNA khỏi sự thuỷ phân của enzyme exonuclease. · Cần cho khởi đầu cho dịch mã. Enzyme thực hiện gắn mũ của tế bào là guanylyl transferase (thêm guauosin 5’-triphosphate) và metyl transferase (thêm nhóm metyl). Các enzyme này nằm trong nhân, nên hầu hết các virus phiên mã trong nhân đều sử dụng. Tuy nhiên các virus phiên mã trong tế bào chất sẽ mã hoá cho enzyme gắn mũ và enzyme metyl hoá của riêng mình. Virus có genome RNA (-) phân đoạn (cúm A) cần có một cơ chế riêng để đoạt lấy mũ từ mRNA của tế bào. Phức hợp protein của virus giúp RNA polymerase của nó gắn vào đầu 5’ của mRNA đã gắn mũ của tế bào, sau đó nhờ hoạt tính endonuclease của phức hợp sẽ cắt một đoạn khoảng 10-20 nucleotide (chứa cả mũ) ra khỏi đầu 5’ của mRNA. Đoạn này sẽ được dùng làm mồi để khởi đầu phiên mã cho virus. Không phải tất cả mRNA virus đều có gắn mũ, mRNA của virus picorna (bại liệt) không có mũ. Đây là virus phiên mã trong tế bào chất, nên không phải vận chuyển ra khỏi nhân. Chúng có thể tiến hành dịch mã theo cơ chế không phụ thuộc mũ, mà nhờ đoạn IRES. f- Gắn đuôi poly (A)Các bản phiên mã đầu tiên của eukaryota và virus được ngắn thêm một loạt gốc adenosin ở đầu 3’ để tạo đuôi poly (A). Sự polyadenin hoá ở đầu 3’ làm tăng tính bền của mRNA tránh sự phân giải của exonuclease và có vai trò nhất định trong khởi đầu dịch mã. Tuy nhiên mRNA của một số virus không gắn đuôi (ví dụ virus reo) nên chức năng kể trên sẽ được thực hiện theo cách khác. Trước vị trí gắn đuôi có một đoạn tín hiệu gắn đuôi dài 20-30bp. Đoạn này ở SV40 là AATAAA, ở HBV là TATAAA (giống như hộp TATA). Khi phiên mã RNA polymerase tiếp tục chạy dọc khuôn, vượt qua tín hiệu gắn đuôi và vị trí gắn đuôi. Sợi RNA mới tổng hợp được cắt tại vị trí gắn đuôi và các gốc adenosin được lần lượt thêm vào nhờ phức hợp protein, trong đó có enzyme gắn đuôi (poly (A) -polymerase). g- Cắt nốiMột số bản phiên mã có chức năng của mRNA nhưng hầu hết còn cần phải chế biến, cắt bỏ intron để tạo mRNA. Hơn nữa từ mRNA, bằng phương thức cắt nối (ghép đoạn này với đoạn kia), có thể tạo ra nhiều loại mRNA khác nhau. Ví dụ bằng phương pháp cắt nối, HIV-1 có thể tạo ra 30 loại mRNA khác nhau. (2) -Dịch mãTham gia vào dịch mã gồm mRNA, tRNA, acid amin và riboxom. Quá trình gồm 3 giai đoạn: Khởi đầu, kéo dài và kết thúc. Ở eukaryota, mRNA là đơn gen (monocistron) có nghĩa là chỉ có một khung đọc, mã hoá cho 1 protein, còn ở prokaryota, mRNA thường là đa gen (polycistron) có nhiều khung đọc và mã hoá cho nhiều protein. Có các vùng không mã hoá, còn gọi là vùng không dịch mã, nằm ở phía đầu 5’ (5’-UTR) và đầu 3’ (3’-UTR). Một khung đọc lớn có thể mã hoá cho một polyprotein lớn sau đó được enzyme phân cắt thành các protein nhỏ với chức năng khác nhau.
Hình 19.4: Dịch mã từ mRNA-monocistron
Một khung đọc mở bắt đầu từ
codon khởi đầu AUG ở đầu 5’ của mRNA và kết thúc tại codon kết thúc (UGA,
UAA, UAG). Dịch mã theo hướng 5’ a- Khởi đầu dịch mãNhư đã nói trên, hầu hết mRNA của eukaryota và virus được gắn mũ và đuôi. Các cấu trúc này có vai trò quan trọng trong dịch mã. - Mũ là nơi bám của các yếu tố khởi đầu dịch mã, eIF (e = eukaryota), tRNA gắn với acid amin (Met-tRNAiMet) và tiểu phần riboxom 40S (hình 6.) - Một protein gọi là protein bám poly (A) bám vào đuôi poly (A). Các protein bám vào đầu sợi RNA có khả năng tương tác và người ta cho rằng sự tương tác này làm cho mRNA truyền tin dẫn đến sự kích thích dịch mã. Các mRNA không có mũ hoặc đuôi có thể truyền tin theo cách khác.
Tiểu phần riboxom 40S di chuyển dọc theo RNA
theo hướng 5’
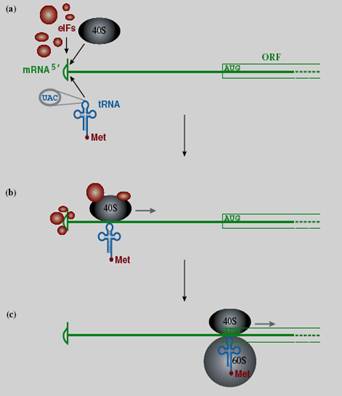
Hình 19.5: Khởi đầu dịch mã trên mRNA có gắn mũ a – Các yếu tố khởi đầu eIF, tiểu phần riboxom 40S và tRNA-methionine bám vào đầu 5’ của mRNA, b - Phức hợp quét từ đầu 5’ theo hướng 3’ của mRNA, c – khi tới codon khởi đầu AUG sẽ được anticodon UAC trên tRNA nhận diện, tiểu phần 60S bám vào và các yếu tố eIF sẽ rời ra. Một số mRNA không có mũ thì sự dịch mã xảy ra theo cơ chế khác. Các eIF thay vì gắn vào mũ, chúng gắn vào vị trí IRES (internal ribosome entry site) gọi là bến đỗ riboxom, đó là đoạn mRNA có cấu trúc bậc 2 ở mức độ cao nằm trước khung đọc, cũng là vị trí gắn của tiểu đơn vị riboxom 40S. Đoạn IRES có ở một số virus RNA, như virus viêm gan C, picorna, đồng thời cũng thấy có ở mRNA của tế bào và mRNA của một số virus khác (ví dụ virus herpes simplex, Sarcoma Rous).
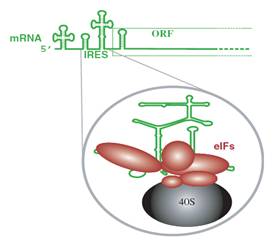
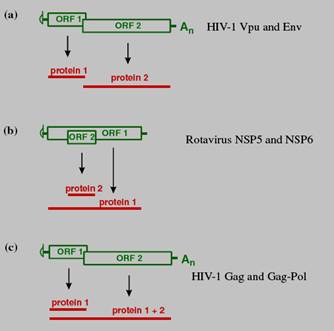
Hình 19.6: Khởi đầu dịch mã trên mRNA không gắn mũ. Tiểu phần roboxom 40S, các eIF bám vào vị trí IRES (bến đỗ riboxom) nằm phía trước khung đọc. b- Dịch mã từ mRNA hai khung đọcHầu hết mRNA của tế bào và của nhiều virus chỉ có 1 khung đọc mở (ORF), nhưng một số mRNA của virus có 2 hoặc nhiều khung đọc, trong số các mRNA bicistron hoặc polycistron này có một số có chức năng là monocistron, nhưng một số mRNA có cấu trúc là bicistron thì cũng có chức năng là bicistron. Sự khác nhau về tỷ lệ dịch mã của hai khung đọc quy định cơ chế biểu hiện 2 gen ở mức độ khác nhau. Ở nhiều mRNA bicistron có khung đọc chồng lớp. Ở trường hợp khác, khung đọc này nằm gọn trong khung đọc kia. - Một cơ chế dùng để đọc khung thứ 2 gồm sự quét bỏ sót (leaky): Tiểu đơn vị riboxom 40S có thể quét bỏ qua codon khởi đầu của khung đọc thứ nhất và bắt đầu dịch mã tại điểm khởi đầu của khung đọc thứ 2. Các khung đọc cho hai protein là khác nhau nên hai protein do chúng mã hoá là khác nhau.
Hình 19.7: Sự dịch mã của mRNA bicistron. a – Riboxom có thể dịch mã tại điểm khởi đầu của ORF-1 hoặc có thể quét qua codon khởi đầu của ORF-2. b – Hai codon khởi đầu là của 2 khung đọc khác nhau và protein được tổng hợp là khác nhau.c – Khung đọc 2 được dịch mã là nhờ sự dịch khung riboxom, tạo ra một dạng kéo dài của protein 1. - Một cơ chế khác để đọc khung đọc thứ hai của mRNA là sự dịch khung (frameshifting) của riboxom: Riboxom nhảy sang khung đọc khác nằm trước điểm kết thúc của khung đọc thứ 1. Do đó nó không bị codon kết thúc của khung đọc thứ nhất nhận ra, mà tiếp tục chạy dọc mRNA để đọc khung đọc thứ nhất kéo dài (hình 19.7c). Sự dịch khung xảy ra khi riboxom di chuyển dọc theo RNA bắt gặp tín hiệu dịch khung (1 trình tự chuyên biệt) có cấu trúc bậc 2, và thường là một pseudoknot (nút giả). c-Cải biến trong và sau dịch mã Trong hoặc sau quá trình dịch mã, protein có thể phải qua một hoặc nhiều lần cải biến, bao gồm glycosyl hoá, acyl hoá hoặc phosphoryl hoá. -Glycosyl hoá Glycosyl hoá là sự gắn thêm nhóm oligosaccarit vào chuỗi polypeptide. Khi một oligosaccarit được gắn vào nhóm –OH của serin hoặc treonin thì gọi là O-glycosyl hoá, còn nếu gắn vào nhóm -NH2 của asparagin thì gọi là N-glycosyl hoá. Protein được tổng hợp trong mạng lưới nội chất hạt, nơi bắt đầu N-glycosyl hoá, sau đó chúng được chuyển tới bộ máy Golgi, nơi hoàn thiện N-glycosuyl hoá nhờ enzyme α-mannosidase I và II, và galactosyl transferase. O-glycosyl hoá được thực hiện trong bộ máy Golgi. Có những virus chỉ cần một loại glycosyl hoá, ví dụ HIV-1 tiến hành N-glycosyl hoá gp120 nhưng cũng có những virus cần cả 2 loại N- và O-glycosyl hoá, ví dụ gC và gD của herpes simplex. Các glycoprotein này được gắn vào vị trí chuyên biệt trên màng sinh chất của tế bào và sẽ là các gai bề mặt sau này của vỏ ngoài. Virus rota không có vỏ ngoài nhưng cũng có glycoprotein. Protein bề mặt VP7 là glycoprotein. Một số glycoprotein của virus là protein không cấu trúc, ví dụ protein NSP4 của virus rota. -Acyl hoá Acyl hoá là sự gắn thêm nhóm acyl (R-CO-) vào phân tử protein. Nhóm acyl thường được gắn là myristyl, trong đó R là CH3-(CH2)12- nhóm myristyl được gắn vào glycin tại đầu N của protein. Hầu như virus không có enzyme N-myristyl transferase để gắn nhóm myristyl, chúng phải sử dụng của tế bào. Protein myristyl hoá gắn vào màng sinh chất, ví dụ protein Gag của hầu hết virus retro. Nếu không được myristyl hoá thì chúng không gắn được vào màng sinh chất và sự lắp ráp virus sẽ không xảy ra. -Phosphoryl hoá Phosphoryl hoá là sự chuyển nhóm phosphate từ một nucleotide, thường là ATP, cho oxy ở nhóm –OH của serin, treonin hoặc tyrosin. Sự chuyển nhóm phosphate nhờ enzyme kinase của tế bào hoặc của virus. Sự phosphoryl hoá có thể làm thay đổi cấu hình, hoạt tính, vị trí và độ bền của protein. Nhiều quá trình của virus đòi hỏi protein phải được phosphoryl hoá. c- Sự vận chuyển các phân tử trong tế bào eukaryota Các phân tử của virus được vận chuyển tới các vị trí nhất định trong tế bào. Genome của virus được đưa vào nhân nhờ trượt theo vi ống và qua lỗ nhân, mRNA virus được vận chuyển từ nhân ra tế bào chất và các protein virus sau khi được tổng hợp có thể được vận chuyển vào các vị trí khác nhau, kể cả vào nhân (Các virus lắp ráp trong nhân). Nhiều protein có một trình tự acid amin giống như mã số vùng của bưu điện, nó quy định điểm đến của protein. Protein đến gắn vào màng có một trình tự tín hiệu – đó là dãy các gốc acid amin kỵ nước. Sự tổng hợp protein bắt đầu tại các riboxom tự do, nhưng khi trình tự tín hiệu được tổng hợp thì nó sẽ hướng dẫn phức hợp polypeptide-riboxom đi vào mạng lưới nội chất và tiếp tục tổng hợp protein tại đó. Vùng mạng lưới nội chất có chứa riboxom vì thế gọi là mạng lưới nội chất sần. Mỗi một loại protein gắn vào màng có một hoặc nhiều trình tự gắn vào màng, gọi là trình tự neo màng (membrane anchor sequences) giàu gốc acid amin kỵ nước. Một số protein loại này có trình tự tín hiệu hoạt động như cái mỏ neo cắm vào màng. Các protein gắn vào màng khác, ví dụ protein vỏ ngoài của HIV-1 sẽ được chuyển qua màng cho đến khi chạm tới trình tự neo và sau đó trình tự tín hiệu sẽ bị enzyme của tế bào loại bỏ. Nhiều protein được tổng hợp trong mạng lưới nội chất hạt sẽ được tạo bọng để tới bộ máy Golgi. Hầu hết protein gắn màng được glycosyl hoá trong các khoang màng này, sau đó được chuyển tới các màng tế bào chất hoặc màng nhân và có thể nẩy chồi qua các màng này. Các tế bào biểu mô có mặt đỉnh (ở phía ngoài) và mặt đáy (ở phía trong), được cấu tạo từ lipid và protein. Khi nhiễm virus có vỏ ngoài vào tế bào biểu mô chúng sẽ nảy chồi qua màng sinh chất một cách giới hạn hoặc qua mặt đỉnh hoặc qua mặt đáy. Ví dụ nếu là VSV (virus chốc mép), chúng sẽ nẩy chồi từ mặt đáy, còn virus cúm A sẽ nảy chồi từ mặt đỉnh. Điều này giải thích vì sao virus cúm nhiễm vào động vật có vú đều khu trú ở đường hô hấp. Nếu virus nhân lên ở trong nhân thì hầu hết (nhưng không phải tất cả) các protein virus phải được đưa vào nhân. Cơ chế vận chuyển liên quan đến sự nhận diện của tín hiệu định vị nhân (NLS-nuclear localization signal) nằm trên protein của tế bào, gọi là importin, sau đó gắn vào các sợi (fibril) mọc ra từ phức hợp lỗ nhân để vận chuyển qua lỗ nhân. (3) - Sao chép genome của virus Đây là bước thứ 5 của chu trình nhân lên. Nhìn chung virus DNA và virus RNA sao chép trực tiếp genome của mình thành RNA. Tuy nhiên một số virus DNA khi sao chép cần qua trung gian RNA và một số virus RNA cần qua trung gian DNA. Các virus DNA tiến hành sao chép trong nhân (trừ virus pox). Các virus có genome nhỏ (virus papilloma) sử dụng DNA polymerase của tế bào, còn virus có genome lớn (ví dụ herpes) thì mã hoá cho enzyme của mình. Các virus RNA tiến hành sao chép trong tế bào chất (trừ virus cúm và retro), sử dụng enzyme do chúng mã hoá. Trong hầu hết trường hợp, sao chép và phiên mã là một. Enzyme dùng cho sao chép cũng là enzyme dùng cho phiên mã. (1) Virus DNA kép sao chép trong nhân theo cơ chế bán bảo tồn như ở tế bào, sử dụng enzyme DNA polymerase phụ thuộc DNA của tế bào. Tuy nhiên virus pox là virus DNA kép, sao chép trong tế bào chất sử dụng enzyme DNA polymerase do chúng mã hoá. (2) Virus DNA đơn (+ hoặc -) tất cả đều sao chép trong nhân, sử dụng DNA polymerase của tế bào và phải qua giai đoạn trung gian tạo sợi DNA kép, gọi là dạng sao chép (RF-replicative form). Tư RF sao chép tạo genome. (3) Virus RNA kép (ví dụ virus rota) luôn có genome phân đoạn, sao chép trong tế bào chất và sử dụng enzyme RNA polymerase phụ thuộc RNA do chúng mã hoá. (4) Virus RNA đơn, (+) khi sao chép phải qua bước tạo RNA (-) trung gian làm khuôn để tổng hợp genome RNA (+). Genome ban đầu được dùng làm mRNA để tổng hợp RNA polymerase. (5) Virus RNA đơn, (-) sao chép trong tế bào chất, sử dụng enzyme RNA polymerase phụ thuộc RNA (RNA-replicase) do chúng mang theo để tổng hợp sợi RNA (+) trung gian làm khuôn tổng hợp genome (-). Virus RNA (-) phân đoạn (ví dụ virus cúm) sao chép trong nhân, sử dụng RNA polymerase phụ thuộc RNA do chúng mang theo. (6) Virus RNA (+) phiên mã ngược (ví dụ HIV), trước hết dùng enzyme phiên mã ngược của virus (DNA polymerase phụ thuộc RNA) để tạo DNA kép trong tế bào chất, sau đó vào nhân gắn vào nhiễm sắc thể của tế bào rồi từ đó sao chép tạo genome RNA nhờ enzyme RNA polymerase phụ thuộc DNA của tế bào. (7) Virus DNA kép phiên mã ngược (ví dụ HBV). muốn sao chép phải qua bước trung gian tạo RNA tiền genome trong nhân, dùng enzyme RNA polymerase phụ thuộc DNA của tế bào. RNA mới sinh ra khỏi nhân làm khuôn để tổng hợp sợi DNA (-) (sợi L) nhờ enzyme phiên mã ngược của virus. Từ sợi DNA (-) làm khuôn tổng hợp sợi DNA (+) tương bù do enzyme của virus mã hoá. a- Khởi đầu sao chépMỗi genome virus có một trình tự đặc biệt, tại đó bắt đầu sao chép acid nucleic của virus. Khi sao chép cần gắn mồi. Đó là phản ứng đầu tiên của một nucleotide với nhóm –OH tại vị trí khởi đầu sự sao chép genome của nhiều virus (ví dụ rota, rhabdo) bắt đầu khi nucleotide đầu tiên của sợi mới bắt cặp với 1 nucleotide trong RNA của virus. Nucleotide đầu tiên hoạt động có hiệu quả như một mồi để sao chép RNA, khi nhóm 3’-OH của nó gắn với nucleotide thứ 2. Một số virus DNA đơn (ví dụ virus parvo) sử dụng cách tự tạo mồi. Đầu 3’ của DNA có các trình tự tương bù, nên có thể gập lại, bắt cặp với nhau tạo đầu 3’-OH thay cho mồi. Để khởi đầu sao chép, nhiều genome DNA và một số genome RNA của virus dùng một phân tử RNA hoặc protein làm mồi. b- RNA mồi và protein mồiSự tổng hợp DNA của tế bào được bắt đầu sau khi một vùng xoắn kép được mở xoắn tạo bong bóng nhờ enzyme helicase và sau khi enzyme primase tổng hợp một đoạn RNA ngắn làm mồi. Cần một mồi cho sợi dẫn đầu và nhiều mồi cho tổng hợp các đoạn Okaseki của sợi muộn (sợi sau). Nucleotide đầu tiên của DNA mới gắn vào 3’-OH của RNA mồi. Một số virus DNA dùng mồi RNA để sao chép genome. Một số (ví dụ virus polymase) dùng primase của tế bào để tổng hợp mồi. Số khác (ví dụ virus herpes và phage T7) lại mã hoá cho primase của riêng mình. Virus retro dùng tRNA của tế bào làm mồi khi ở ngoài tế bào chất, nhưng khi cài xen genome của mình vào nhiễm sắc thể của tế bào nên để sao chép, chúng dùng mồi do primase của tế bào tổng hợp. Một số virus động vật sử dụng protein làm mồi, trong đó có virus DNA như virus adeno và virus RNA như picorna. Nhóm 3’-OH của serin hoặc tyrosin trong protein sẽ gắn với nucleotide sợi mới. Virus hepDNAa (ví dụ HBV) là virus DNA kép, dùng mồi là protein để khởi đầu tổng hợp sợi DNA (-) và mồi RNA để khởi đầu tổng hợp DNA (+). Mồi protein và mồi RNA của virus hepDNAa không bị cắt bỏ sau khi vai trò của chúng đã hoàn tất mà vẫn được dính vào đầu 5’ của genome. c- Sao chép DNAMỗi DNA virus có ít nhất một trình tự chuyên biệt để bắt đầu sao chép gọi là trình tự khởi đầu (Ori). Các protein khởi đầu sao chép DNA bám vào vị trí này bao gồm: - Helicase bám vào vị trí để tháo xoắn. - Một số protein bám sợi DNA đơn, giữ cho 2 sợi không bắt cặp lại với nhau. - Một DNA polymerase. Về cơ bản quá trình sao chép DNA của virus giống như của tế bào. Ở vi khuẩn, số enzyme tham gia ít hơn so với ở eukaryota. Ví dụ helicase-primase của phage T7 (ở E.coli) chỉ là 1 phân tử, trong khi của virus herpes simplex (ở tế bào động vật) là một phức hợp gồm 3 loại protein. Sự tổng hợp DNA diễn ra gần chạc sao chép. Một trong 2 sợi là sợi dẫn đầu, sợi còn lại là sợi muộn, được tổng hợp thành các đoạn Okaseki sau đó nối lại với nhau nhờ DNA-ligase. Sợi DNA kép mới tạo thành có chứa một mạch của sợi mẹ. Cách sao chép này gọi là bán bảo tồn. Ngược lại với nó là sao chép bảo tồn xảy ra ở một số virus. Một số genome DNA là phân tử dạng thẳng trong khi một số khác lại là dạng khép vòng. Một số phân tử dạng thẳng khi sao chép lại được khép vòng, cho nên nhiều genome virus được sao chép như là một phân tử vòng tròn. Từ đây có 2 phương thức sao chép. - Sao chép theo cơ chế theta hay dạng mắt. Từ vị trí Ori tạo ra 2 chạc ba sao chép. Sao chép cùng lúc theo 2 chiều thuận nghịch kim đồng hồ. - Sao chép theo cơ chế xích ma (d). Phân tử DNA kép dạng vòng gồm sợi ngoài (+) và sợi trong (-). Sợi ngoài bị cắt đứt ở liên kết photphodiete tạo ra đầu 3’-OH tự do (gọi là điểm sinh trưởng) Sợi trong xoay được dùng làm khuôn. Các nucleotide nối vào đầu 3’-OH để tạo ra 1 sợi DNA mới. Từ sợi DNA mới này lại được gắn mồi tổng hợp mạch bổ sung tạo DNA kép. Một số virus lúc mới nhiễm sao chép theo cơ chế theta nhưng ở giai đoạn sau lại theo cơ chế xích ma (ví dụ phage fx174). Một số virus DNA như virus herpes và phage T4, kết quả sao chép tạo ra phân tử DNA rất lớn gọi là phân tử trùng lặp (concateme). Mỗi concateme cấu tạo gồm nhiều bản sao genome nối với nhau. Trước khi lắp ráp vào virion concateme sẽ được phân cắt thành các phân tử có kích thước và trình tự của genome. (4) -Lắp ráp Lắp ráp là sự tự kết nối các thành phần virus để tạo ra virion hoàn chỉnh đòi hỏi phải có cấu trúc bền vững, tồn tại được trong môi trường như là một thực thể có khả năng gây nhiễm, tuy nhiên cũng đòi hỏi khi vào trong tế bào cấu trúc này phải không bền vững thì mới có thể giải phóng dễ dàng genome vào tế bào chất. Do vậy virion phải có cơ chế đóng mở kiểu “công-tắc” để có thể biến đổi từ trạng thái bền vững sang trạng thái không bền vững. Công tắc này liên quan đến việc gắn vào receptor hoặc sự thay đổi pH trong endosome. Khi số lượng genome và protein cấu trúc được tích luỹ đến ngưỡng thì chúng sẽ tiến hành lắp ráp tạo nucleocapsid. 1. Virus có cấu trúc dạng xoắn: Đối với virus RNA đơn có cấu trúc dạng xoắn, lúc đầu một số phân tử protein cấu trúc sẽ bám theo chiều xoắn của genome RNA, sau đó các phân tử khác lần lượt bám theo cho đến khi phủ hết RNA. 2. Virus có cấu trúc dạng khối đa diện: Trước hết cần phải lắp ráp một cấu trúc rỗng hình cầu gọi là procapsid. Genome virus chui vào procapsid, sau đó cải biến từ cấu trúc hình cầu sang hình khối đa diện; ví dụ các virus adeno, picorna thực hiện cải biến bằng cách cắt bớt 1 hoặc nhiều protein cấu trúc. Genome chui vào trong procapsid qua kênh nằm ở vị trí mà sau này sẽ là đỉnh của khối đa diện. Bất kỳ enzyme nào tham gia vào đóng gói genome cũng nằm ở vị trí này. Ở tế bào thực khuẩn cũng như vậy, trước hết cũng phải tạo một “tiền đầu” (prohead) sau đó genome chui qua 1 cái cổng nằm ở một đỉnh. Vị trí này cũng có chức năng nối với đuôi. a- Đóng gói genomeTrong tế bào có rất nhiều acid nucleic, của cả virus và tế bào. Vậy làm thế nào để genome của virus lại được lựa chon và lắp ráp chứ không phải của tế bào? Sở dĩ như vậy vì virus có một protein chuyên biệt, nhận diện tín hiệu đóng gói, nằm ở vùng có cấu trúc bậc hai của genome. Hầu hết virus có genome sợi đơn có thể đóng gói hoặc sợi dương hoặc sợi âm, nên tín hiệu đóng gói phải có duy nhất ở sợi cần được đóng gói. Genome được nén trong thể tích nhỏ. Virus DNA kép có kích thước lớn như virus herpes, đóng gói genome chặt đến nỗi tạo áp suất lớn gấp 10 lần so với áp suất trong chai rượu sâm – banh. b- Cơ chế lắp ráp Trước đây Fraenkel-Conrat đã tách genome ra khỏi capsid của virus đốm thuốc lá, sau đó lại lắp ráp chúng với nhau trong điều kiện pH và sự có mặt của một số ion nhất định, để tạo virus hoàn chỉnh. Với các virus đơn giản, chỉ chứa 1 acid nucleic và một số ít loại protein thì có thể tự lắp ráp một cách đơn giản như trên. Nhưng với các virus phức tạp, như virus herpes và phage có đuôi, thì không thể tái lắp ráp như vậy. Chúng cần phải được lắp ráp trực tiếp trong môi trường của tế bào nhiễm. Khi lắp ráp cần phải có mặt tạm thời 1 protein dùng làm giàn giáo. Các protein cấu trúc theo đó mà lắp vào để tạo capsid. Khi công việc hoàn tất, protein giàn giáo bị loại bỏ khỏi virion hoặc bằng enzyme phân giải hoặc giữ lại để tái sử dụng. c- Sự tạo màng virion Vỏ ngoài virus có thể được tạo thành theo 1 trong 2 cơ chế: Cải biến màng sinh chất của tế bào rồi nảy chồi ra ngoài hoặc tự tổng hợp màng mới bao quanh nucleocapsid. -Cải biến màng sinh chất Vỏ ngoài của virus thường có nguồn gốc từ màng sinh chất, được cuốn theo khi virus nảy chồi. Vùng màng mà virus sẽ nảy chồi được đính trước 1 hoặc nhiều loại protein đặc hiệu của virus, thường là glycoprotein, các protein này ngâm trong lớp lipid kép. Protein M của virus (các phân tử protein này gắn với nhau tạo thành màng đệm – M) tập trung nhiều ở vùng màng, có ái lực với nhau và đẩy protein tế bào ra khỏi màng. Đôi khi protein tế bào không bị đẩy ra hết nên chúng có thể tham gia vào thành phần vỏ ngoài. Ví dụ vỏ ngoài của virus HIV-1 có chứa protein MHC-II (phức hợp hoà hợp mô chính) của tế bào. Trước khi nảy chồi, protein M tới gắn vào phần đuôi nằm trong tế bào chất của glycoprotein xuyên màng, sau đó nucleocapsid tiến đến bám vào màng M. Nucleocapsid khi nảy chồi sẽ cuốn theo màng tế bào chất và màng M để tạo vỏ ngoài. Không phải tất cả các virus đều có màng M. Ví dụ ở trường hợp virus sốt vàng thì bề mặt nucleocapsid sẽ gắn trực tiếp vào đuôi glycoprotein trong màng. -Tổng hợp mới màng virus Chỉ có một số ít virus tạo màng lipid muộn trong quá trình nhân lên. Màng này có thể tham gia hình thành vỏ ngoài (ví dụ virus pox) hoặc nằm trong nhân tạo 1 lớp phía mặt dưới của capsid (ví dụ virus irido). Khi nhân lên virus baculo tạo ra 2 loại virion có vỏ ngoài: Loại thứ nhất có chức năng lây nhiễm sang tế bào khác trong vật chủ. Loại này có được vỏ ngoài là do nảy chồi qua màng sinh chất. Loại virion thứ hai có chức năng gây nhiễm vào vật chủ mới, vỏ ngoài của nó bao quanh nucleocapsid nằm trong nhân dẫn đến các virion hợp nhất với nhau trong một bọc gọi là thể bọc (occlusion body). Thể bọc giúp virus tránh tác động của môi trường bên ngoài, nên virus có thể duy trì rất lâu ngoài tế bào sống. (5) -Giải phóng virus khỏi tế bào Đây là giai đoạn cuối cùng của chu trình nhân lên. Nhiều virus được giải phóng khi tế bào bị nổ tung, do thành tế bào bị phân giải, cộng với áp lực lớn trong tế bào. Ví dụ nhiều phage, peptidoglycan bị phân giã dẫn đến làm vỡ tế bào. Số khác lại có khả năng tổng hợp protein ức chế enzyme tham gia vào tổng hợp thành tế bào, làm cho thành tế bào yếu đi nên dễ bị vỡ. Số lượng virus được tạo thành qua mỗi mẻ là rất lớn. Trong tế bào E.coli có kích thước nhỏ bé, phage T4 có kích thước lớn tạo ra được 200 virion. Virus picorna có kích thước rất nhỏ khi nhiễm vào tế bào động vật có kích thước lớn đã tạo ra đến 100.000 virion. Một số virus không làm tan tế bào mà chỉ chui ra từ từ theo lối nảy chồi, số khác chui ra thông qua việc tạo thành túi hay bọng từ màng lưới nội chất hoặc bộ máy Golgi. Màng bọng dung hợp với màng tế bào chất và đẩy virus ra ngoài. Ở các trường hợp này, tế bào vẫn còn sống thêm một thời gian nữa. 19.1.3.2.Phiên mã và dịch mã ở Prokaryota 19.1.3.2.1. Phiên mã ở Prokaryota Sự phiên mã ở các tế bào prokaryota có những đặc điểm sau: - Chỉ cần một loại RNA polymerase để tổng hợp tất cả các loại RNA. - mRNA là đa gen (polycistron) có nhiều khung đọc, tất cả các khung đọc đều có thể được dịch mã cùng một lúc. - mRNA không gắn mũ và đuôi poly (A). - Phiên mã được thực hiện khi RNA-polymerase bám vào promoter. Sự tổng hợp bắt đầu từ điểm xuất phát, thường là TAC, nằm sau điểm bám 7-8 base nằm phía đầu 3’ của khuôn. - Phiên mã tiếp tục cho đến khi đọc qua trình tự kết thúc. Khi kết thúc, RNA polymerase và mRNA rời khỏi mạch khuôn. - Quá trình phiên mã và dịch mã xảy ra đồng thời. RNA-polymerase của E.coli là một phức hợp gồm 5 tiểu đơn vị là ααbb’ và d. d (xích ma) có thể tách ra khỏi enzyme lõi. ααbb’ (enzyme lõi) là yếu tố xác định tính đặc hiêu của promoter. Promoter là vùng khởi động. Nếu ký hiệu base đầu tiên phiên mã thành mRNA (thường là adenin) là +1 thì các base nằm phía trước theo hướng ngược chiều phiên mã sẽ ký hiệu là (-). Promoter gồm 2 trình tự một trình tự là TATAAT cách điểm khởi đầu 5-8 base và có base trung tâm là -10. Trình tự này gọi là hộp TATA hay Pridnow và một trình tự nữa là TTGACA, có base trung tâm là -35 (nằm trước điểm khởi đầu 35 bp). Vùng -35 tham gia vào việc gắn RNA polymerase
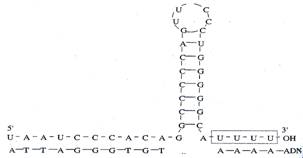
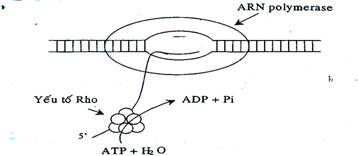
Cụ thể là ngay trước vùng -35 của một số promoter còn có thêm yếu tố UP để cho tiểu đơn vị a của RNA polymerase nhận diện và tăng cường sự bám của enzyme. Khi khởi đầu phiên mã hoàn thành, yếu tố a sẽ rời ra để tái sử dụng. Phiên mã kết thúc sau khi một trình tự kết thúc được phiên mã. Sự kết thúc có thể theo cơ chế phụ thuộc hay không phụ thuộc yếu tố r (rho). - Kết thúc phiên mã không phụ thuộc rho được đặc trưng bởi trình tự giàu G-C trên DNA. Trình tự này đối xứng 2 bên, có thêm 5 hoặc 6 adenin kèm theo. RNA được phiên mã từ trình tự này có thể tạo ra cấu trúc nút vòng có cuống (stem loop) và cấu trúc nối A-U hình thành trong sợi lai DNA-RNA (DNA khuôn và RNA mới sinh). Đoạn lặp lại oligo A với oligo U bắt cặp không bền vững nên RNA có thể tách khỏi DNA khuôn. Sau đó sợi DNA kép được hình thành trong “bong bóng phiên mã”. Lõi của enzyme RNA polymerase có ái lực thấp với DNA kép nên được tách ra.
Hình 19.8: Cấu trúc bậc 2 ở đầu 3’ của RNA ở operon E.coli. - Kết thúc phiên mã phụ thuộc yếu tố rho cần phải có yếu tố rho. Đó là protein gồm có 6 tiểu đơn vị có ái lực cao với RNA đơn, có hoạt tính helicase và ATP-ase để tháo xoắn sợi lai DNA-RNA. Khi bám vào RNA, yếu tố rho sẽ phân giải ATP. Năng lượng được giải phóng giúp nó chuyển dọc sợi RNA mới sinh tới bong bóng phiên mã, sau đó yếu tố rho tách đôi DNA-RNA và giải phóng RNA. Gen của virus và vi khuẩn ít khi có intron. Một số phage dùng RNA pol phụ thuộc DNA của tế bào vật chủ để phiên mã, trong khi số khác lại tự tổng hợp enzyme này cho riêng mình.
Hình 19.9: Mô hình kết thúc phiên mã phụ thuộc yếu tố Rho ở E. Coli 19.1.3.2.2. Dịch mã ở prokaryota Dịch mã ở vi khuẩn có các đặc điểm sau: - Dịch mã có thể được bắt đầu trước khi kết thúc phiên mã. Do không có nhân nên phiên mã và dịch mã xảy ra đồng thời. - Riboxom 70S gồm 2 tiểu phần là 50S và 30S. - mRNA không có mũ nhưng có trình tự SD (Shine DalgRNAo) nằm trước vị trí khởi đầu dịch mã AUG và bắt cặp với đoạn 3’RNA riboxom 16S trong tiểu phần 30S. - Methionine của tRNA đầu tiên thường được metyl hoá (fMet-tRNAiMet). - Chỉ cần một lượng rất ít các yếu tố khởi đầu. - mRNA có nhiều khung đọc (polycistron). Tất cả các khung đọc đều được dịch mã cùng lúc.
Hình 19.10: Quá trình nhân lên của virus trong tế bào vật chủ
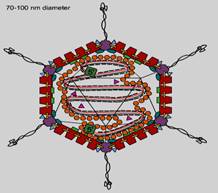
Hình 19.11: Sự xâm nhập của thực khuẩn thể vào tế bào vi khuẩn 19.2. GIỚI THIỆU TÓM TẮT QUÁ TRÌNH NHÂN LÊN CỦA MỘT SỐ VIRUS ĐIỂN HÌNH 19.2.1. Virus DNA 4 họ đại diện là: Adenoviridae, Herpesviridae, Papovaviridae và Parvoviridae. 19.2.1.1. Adenoviridae Họ này gồm các chi: - Mastadenovirus, gồm 49 serotyp gây bệnh cho người như viêm phổi, viêm mũi họng, viêm kết mạc, một số gây ung thư ở chuột. - Aviadenovirus, nhiễm ở chim, gia cầm. - Virus chứa genome DNA kép. - Virion dạng khối đa diện, trên mỗi đỉnh có sợi lông dài. - Không có vỏ ngoài. a-Cấu trúc
- Đường kính trung bình của virion 70-100nm - Capsid chứa 7 protein: Protein II nằm trên vỏ capsid. Protein III hay protein penton, nằm ở đỉnh khối đa diện. Protein IIIa nằm trên vỏ capsid. Protein IV sợi đỉnh. Protein VI nằm ở gần đỉnh. Protein VIII nằm trên cạnh tam giác của khống đa diện. Protein IX nằm gần đỉnh. - Lõi chứa các protein. Protein V nối DNA với protein II (penton). Protein VII bao quanh sợi DNA. Protein X nằm trên bề mặt tam giác của capsid. Protein 55 kDa gắn ở đầu chuỗi gọi tắt là protein đầu chuỗi (TP). - Ở đầu chuỗi DNA có các trình tự lặp lại đảo chiều.
b- Hấp phụ và xâm nhập - Sợi đỉnh của virus gắn vào thụ thể (CAR) dành cho kháng thể của virus coxsackia và adeno. - Intergrin gắn vào trình tự RGD trong protein III (penton) để giúp virus xâm nhập vào tế bào theo lối nhập bào, tạo endosome. - Virus thoát khỏi endosome nhờ pH trong endosome thay đổi, bị vỡ ra giải phóng nucleocapsid. - Vi ống gắn với hexon và vận chuyển virion vào gần lỗ nhân. Vỏ capsid bị phân giải, DNA được giải phóng và vào nhân qua lỗ nhân. Protein 55 kDa ở đầu 5’ bám vào màng matrix của nhân. c- Phiên mã và dịch mã sớm Genome được tách thành sợi R và L (phải và trái) không gọi là sợi (+) và sợi (-) vì cả 2 sợi đều có khung đọc. · Cả 2 sợi đều phiên mã tạo mRNA. · Sợi R có các gen E1A, E1B, các gen cấu trúc, gen E3. · Sợi L có các gen E2A, E2B và E4. - RNA polymerase II phiên mã tạo mRNA của virus có mũ ở đầu 5’ và đuôi poly (A) ở đầu 3’. · mRNA E1 dịch mã tạo protein E1. · Các gen sớm trung gian E1A tham gia vào hoạt hoá trans các gen virus. - Protein E1 gắn vào protein điều hoà của vật chủ, gắn vào hộp TATA, điều hoà ngược pha S, hoạt hoá protein p53. · Protein E1B ức chế sự chết theo lập trình (apoptosis) của tế bào. · E1 gắn pRB (protein ung thư võng mạc). - Phiên mã sợi L để tạo mRNA cho E4 sau đó dịch mã để tạo protein E4. · E4 điều hoà ngược các gen virus, tham gia vào vận chuyển mRNA và vào tổng hợp DNA. - Phiên mã mRNA E2, tổng hợp 3 protein quan trọng cần cho tổng hợp DNA. - Phiên mã mRNA E3, dịch mã tạo ra protein E3 để cải biến đáp ứng miễn dịch của vật chủ ức chế apoptosis. d-Sao chép Sao chép DNA tiến hành khi tế bào bước vào pha S và khi E2 được tích luỹ. - Các đoạn lặp đảo chiều ở 2 đầu dùng làm điểm khởi đầu sao chép. - Có một protein 80 kDa (protein đầu chuỗi) và một DNA polymerase. Từ vùng E2 tạo 1 heterodimer để bắt đầu sao chép. · Cả 2 sợi của genome đều có thể bị thế chỗ do virus thực hiện sao chép theo cơ chế bán bảo thủ. · Sợi mẹ có cấu trúc cán chảo (panhandle) do có các đoạn đầu lặp đảo chiều và tiến hành phiên mã. · Protein đầu chuỗi 80 kDa bị cắt bớt để tạo thành protein 55 kDa. e- Phiên mã và dịch mã muộn - Sau sao chép, sự vận chuyển mRNA của vật chủ bị ức chế. · Phức hợp E4.E1B ngăn cản sự vận chuyển mRNA của vật chủ và tăng cường sự vận chuyển mRNA của virus ra khỏi nhân. · Sản phẩm của gen sớm sau hoạt hoá trans các gen muộn. - Sự cắt nối mRNA và vị trí gắn đuôi poly (A) khác nhau tạo nên các mRNA khác nhau. f- Lắp ráp và giải phóng - Penton và hexon đầu tiên được lắp ráp trong tế bào chất, sau đó tập hợp trong nhân để lắp ráp thành capsid. - DNA chui vào capsid nhờ phân tử protein đóng gói. Nếu DNA không chui được cào capsid, nó sẽ tách ra ở ngoài. Virion hoàn chỉnh ra khỏi nhân rồi toát ra ngoài. 19.2.1.2. Herpesviridae Họ này gồm 3 chi: (1) Alpha herpesvirus - HHV-1 (human herpesvirus-1) gây chốc mép. - HHV-2, còn gọi là herpes sinh dục gây bệnh đường sinh dục. - HHV-3 gây bệnh thuỷ đậu, zona. - Simian herpesB gây bệnh ở khí. (2) Beta herpesvirus - HHV-4 virus Epstein-Barr gây ung thư vòm họng. - HHV-5 virus cự bào (CMV-cytomegalovirus). - HHV-6 gây bệnh ban (ban đào). - HHV-7 không biểu hiện triệu chứng. - HHV-8 Sarcoma Kaposi ở người.
(3) Gamma herpesvirus a- Cấu trúc
- Virus có vỏ ngoài, trên bề mặt có các glycoprotein, glycoprotein bề mặt gC, gD, gB, gE, gH, gI, gK, gL, gM. Vỏ ngoài chứa lipid đường kính 150-200nm. - Phía trong vỏ ngoài là lớp protein vô định hình (tegument) – vùng hạt chứa protein dạng cầu. - Capsid có kích thước trung bình (100nm), dạng khối đa diện với 162 capsome. - Lõi là DNA kép, dạng thẳng là một trong những genome lớn nhất của virus (240 kb). b- Hấp phụ và xâm nhập - Protein bề mặt gC của virus bám vào thụ thể bề mặt của tế bào. + Một số chất trung gian xâm nhập (HVEM-herpesvirus entry mediators) giúp virus gắn dặc hiệu vào glycoprotein gD. + Khi dung hợp với màng tế bào cần có thêm sự tham gia của các glycoprotein bề mặt khác gB, gD, gE và gI. - Virus vào tế bào theo lối nhập bào. - Virus tiếp cận nhân, chỉ có DNA và protein tegument mới qua lỗ nhân. c- Phiên mã và dịch mã sớm - Genome virus khép vòng nhưng không liên kết với protein chromatin. - Một phần protein tegument của virus herpes simplex vẫn còn giữ lại ở tế bào chất sẽ ngăn cản sự tổng hợp protein của tế bào chủ nhờ phân huỷ polyriboxom và RNA của tế bào. + Protein tugument, trong đó có a-TIF (yếu tố cảm ứng trans của gen a) tiến hành hoạt hoá trans và tăng cường tổng hợp mRNA sớm ngay. + 5 gen a sớm ngay (khoảng 10%) không đòi hỏi tổng hợp protein mà là nguồn dự trữ cho sản phẩm gen a. - RNA pol II phiên mã các gen a còn lại tạo mRNA để tổng hợp protein a tham gia vào sự điều hoà biểu hiện gen của virus herpes. - Các gen b (gen sớm sau) được phiên mã ở mức độ thấp nếu không có sản phẩm của gen a. - Gen b1 cần sản phẩm của gen a và được phiên mã trước tiên, tạo mRNA để tổng hợp ICP-6 (b16) là ribonucleotide reductase. ICP-18 (b18) là protein bám DNA. - b2 là gen sớm sau phụ thuộc vào gen a4 và được phiên mã sau gen b1. - Một số protein b2 ngăn cản sự tổng hợp protein a. Các sản phẩm của gen b đạt cao nhất sau 5-7 giờ nhiễm. d- Sao chép genome - Trước khi tổng hợp DNA đã có một số protein cấu trúc g1. - Virus sử dụng nhiều enzyme như helicase, primase và polymerase để tiến hành sao chép genome theo cơ chế vòng tròn xoay. - Sao chép bắt đầu tại 3 điểm khởi đầu và nhiều đoạn RNA không được loại bỏ. Khi DNA sao chép tại nhiều điểm khởi đầu sao chép, lúc đầu được sợi đơn rất dài, sau đó tổng hợp sợi bổ sung. Đây là sợi trùng lặp (concateme) gồm nhiều genome nối với nhau. Enzyme sẽ cắt để được phân tử có kích thước và trình tự của genome. e- Phiên mã và dịch mã muộn - Các protein b hoạt hoá gen g1. Một số protein g1 được tạo thành trước khi tổng hợp DNA. - Các gen g mã hoá cho các protein muộn - tức protein cấu trúc, tích luỹ trong nhân và ở màng nhân, ví dụ các capsome dạng ống, các protein bề mặt vỏ ngoài. - Các protein g ức chế sự tổng hợp sản phẩm gen b.
Hình 19.12: Sự điều hoà biểu hiện gen của virus herpes.
Gen a được hoạt hoá bởi protein a-TIF do gen b mã hoá. Protein a tự điều hoà biểu hiện của chính mình và hoạt hoá gen b. Protein a và b hiệp đồng hoạt hoá gen g, proetin g mà tham gia lắp ráp tạo virion vừa điều hoà hoạt động của gen a.
Ghi chú: f- Lắp ráp và giải phóng Protein tạo capsid gắn với trình tự “a” nằm trên DNA kép, cắt chúng ra khỏi sợi trùng lặp, tạo capsid, qua màng nhân. Màng nhân bọc capsid sau đó dung hợp với màng sinh chất để ra ngoài. 19.2.1.3. Papovaviridae Gồm các chi: Papilomavirus gây bệnh mụn cóc, ung thư cổ tử cung. Polymevirus gây viêm não. Virus tạo bọt SV40. a- Cấu trúc
Capsid hình khối đa: diện, đường kính 45-55nm, không có vỏ ngoài, được tạo thành từ 72 capsome. Capsome được cấu tạo từ các protein VP1, VP2 và VP3. VP1 là protein lớn của capsid. VP2 là protein nhỏ của capsid. VP3 là protein capsid gắn DNA. Lõi là DNA kép, khép vòng. b- Hấp phụ và xâm nhập - Virus vào tế bào biểu mô theo cơ chế nhập bào. Sự hấp phụ thay đổi tuỳ loại virus, nhưng có thể cần phân tử MHC-I, yếu tố sinh trưởng hoặc acid sialic. thụ thể của HPV-6 là integrin, của HPV-11 là heparin sulphat. - Virus nằm trong endosome và cởi vỏ ở tế bào chất, sau đó genome DNA kép, khép vòng vào nhân qua lỗ nhân. c- Phiên mã và dịch mã sớm Phiên mã nhờ RNA polymerase II của tế bào để được pre-mRNA (mRNA tiền chất). Pre-mRNA cắt nối tạo 2 mRNA, nhưng có cùng mũ ở đầu 5’ và đuôi poly (A) ở đầu 3’. Có tên gọi là kháng nguyên T lớn và kháng nguyên T nhỏ. Virus SV40 có 2 kháng nguyên T trong khi virus polyoma có 3 kháng nguyên T (thêm kháng nguyên T trung bình). d- Sao chép genome - Sự tích luỹ kháng nguyên T lớn có tác dụng chuyển tế bào vào pha S, bởi vì virus phụ thuộc vào bộ máy sao chép của tế bào. - DNA của virus chuyển thành một nhiễm sắc thể nhỏ (minichromosome). - Kháng nguyên T lớn bám vào DNA và hoạt hoạt động như enzyme helicase và ngăn chặn sự phiên mã sớm bằng cách phong bế điểm khởi đầu. - Sự sao chép theo cơ chế theta tạo ra 2 genome giống hệt nhau. e- Phiên mã và dịch mã muộn - Sự chuyển vị trí khởi đầu sau sao chép DNA cho phép bắt đầu tổng hợp mRNA muộn. - Nhờ phương thức cắt nối lựa chọn (alternative splicing) mà từ 1 bản phiên mã tạo ra được 3 mRNA có đoạn dẫn đầu không dịch mã giống nhau. + mRNA VP1 tổng hợp protein Vp1. Các riboxom đôi khi bắt đầu dịch mã tại điểm khởi đầu dịch mã thay đổi để tạo ra VP3. + mRNA Vp2/VP3 tổng hợp các protein VP2/VP3.
f- Lắp ráp và giải phóng - VP2 và VP3 gắn vào VP1 và tín hiệu định vị nhân trên VP1 tạo thành phức hợp tích luỹ trong nhân. - Các protein cấu trúc tự động lắp ráp với nhau tạo thành capsid rỗng. DNA loại bỏ histon-1 trước khi chui vào capsid. - Nucleocapsid rời khỏi nhân, tạo bọng với màng lưới nội chất nhẵn để ra khỏi bề mặt đỉnh của tế bào biểu mô nhờ dung hợp với màng tế bào chất. 19.2.1.4. Parvoviridae Gồm các chi: - Parvovirus gây bệnh ở động vật gậm nhấm, bệnh ở chó, lợn, bệnh mắt bạch tạng cầu hạt ở mèo,... - Erytrovirus gây bệnh ban đỏ B19. - Dependovirus khác với 2 chi trên là các virus tự lập (autonomous) có thể tự nhân lên trong các tế bào chủ thích hợp. Dependovirus là virus khiếm khuyết, chúng chỉ có thể nhân lên khi có sự hỗ trợ của virus đồng nhiễm, ví dụ virus adeno. Virus đi kèm adeno. Virus đi kèm adeno ở chim. Virus đi lèm adeno ở chó. a- Cấu trúc
- Capsid có kích thước rất nhỏ (20nm), dạng khối đa diện, gồm 32 capsome với 60 tiểu đơn vị VP (viral protein) là VP1, VP2 (chiếm 90% tiểu đơn vị protein) và VP3. - Lõi: DNA đơn, dạng thẳng. Hầu hết là DNA (-), ở hai đầu có đoạn palindrom (đoạn DNA mạch kép có trình tự nucleotide trên mỗi sợi giống nhau nhưng trái chiều nhau) tạo thành các nút kẹp tóc, có một đầu 3’-OH thay cho mồi. b- Xâm nhập và cởi vỏ - Virus B19 gắn vào thụ thể là kháng nguyên P trên bề mặt hồng cầu. Các cá thể không có thụ thể này sẽ không bị nhiễm. Cách xâm nhập và cởi vỏ còn chưa thật rõ. - DNA đơn vận chuyển vào nhân. c- Phiên mã và dịch mã sớm - RNA polymerase của tế bào tổng hợp mRNA của đoạn nằm trước hộp TATA để tổng hợp protein không cấu trúc, tức là protein điều hoà. - Cắt nối mRNA, tạo ra các bản sao RNA, gắn mũ và đuôi, được mRNA ra khỏi nhân để tổng hợp protein điều hoà NS1. - Tương tự như vậy, tiếp tục cắt nối trong nhân để tạo mRNA dùng cho tổng hợp protein điều hoà NS2. - Các protein không cấu trúc NS1 và NS2 vận chuyển vào nhân để tham gia sao chép DNA. d- Sao chép genome - Sự sao chép genome chỉ được tiến hành khi tế bào chủ ở pha S của chu kỳ tế bào. - Đối với virus phụ thuộc adeno (AAV) đòi hỏi phải đồng nhiễm với virus adeno để kích tế bào bước vào pha S. - Để sao chép, cần có DNA polymerase và một số enzyme của tế bào. Sao chép dựa trên mô hình kẹp tóc lăn cải biến. Sao chép bắt đầu từ đầu 3’ của nút kẹp tóc, được dùng thay cho mồi, để tổng hợp sợi DNA (+) bổ sung, kéo dài đến khi đạt chiều dài genome. - Protein NS1 cắt và liên kết cộng hoá trị với genome, tạo ra điểm đứt trong mạch ban đầu cho phép nút kẹp tóc duỗi thẳng. Đoạn mới duỗi dùng làm khuôn để tổng hợp đoạn DNA (+) bổ sung. - Chu kỳ tiếp tục lặp lại tạo phân tử DNA trùng lặp (concateme) rất lớn. - Protein NS1 gắn vào mỗi trình tự genome để cắt, tạo genome hoàm chỉnh. e- Phiên mã và dịch mã muộn - Nhờ cắt nối lựa chọn phân tử mRNA, mà tạo được 2 phân tử mRNA cho tổng hợp 2 protein cấu trúc khác nhau là VP1 và VP2. - VP2 lại phân cắt trong capsid nhờ enzyme để tạo VP3. f- Lắp ráp và giải phóng - NS1 đóng gói DNA virus ở trong nhân. - Các protein cấu trúc (VP) vào nhân qua lỗ nhân tạo capsid. - Cơ chế chui ra của virus còn chưa rõ.
19.2.2. Virus RNA Virus có genome RNA, bao gồm RNA đơn (+), RNA đơn (-) và RNA kép, có số lượng họ rất lớn. Sau đây xin giới thiệu các họ quan trọng nhất. 19.2.2.1. Virus RNA đơn, dương Gồm một số họ đại diện: - Coronaviridae. - Flaviviridae. - Picornaviridae. - Retroviridae. - Togaviridae. 19.2.2.1.1. Coronaviridae Có 1 chi là chi coronavirus gây bệnh: - Viêm đường hô hấp cấp nặng (SARS). - Coronavirus ở người (HCoV) gây bệnh đường hô hấp. - Viêm phế quản ở chim (IBV). - Viêm gan chuột (MHV). - Bệnh mào xanh ở gà Tây.
a-Cấu trúc
Virus có kích thước trung bình (80-100nm) Vỏ ngoài có các protein: - Protein xuyên màng. - Protein hemagglutinin esterase. - Protein gai, hình dùi cui, gọi là teplome, nên tạo diềm quanh virus. Khi quan sát dưới kính hiển vi, diềm có hình ảnh vầng hào quang (corona tiếng Latinh la hào quang). Nucleocapsid dạng xoắn, mềm mại. Protein nucleocapsid liên kết với genome. Lõi Genome là RNA đơn, (+). b- Hấp phụ và xâm nhập - Protein gai giúp virus bám vào một loạt glycoprotein bề mặt của tế bào. - Virus xâm nhập theo 2 cách: dụng hợp với màng sinh chất và nhập bào. - Sự cởi vỏ và giải phóng RNA vào tế bào chất còn chưa rõ. c- Dịch mã sớm Genome RNA, (+) có chức năng mRNA, tiến hành dịch mã sớm tạo protein còn chưa rõ chức năng. d- Phiên mã - Phiên mã tạo sợi khuôn RNA (-), từ đó tổng hợp rất nhiều mRNA khác nhau. - Phiên mã tạo sợi RNA (-) không liên tục, rồi từ đó tổng hợp rất nhiều mRNA. e- Tổng hợp protein - Từ các mRNA tổng hợp nhiều loại protein khác nhau, như glycoprotein gai, protein xuyên màng, protein heamagglutinin, protein cấu trúc. f- Lắp ráp và giải phóng - RNA genome cuộn lại, gắn với protein nucleocapsid. - Các glycoprotein gắn vào màng bộ máy Golgi, nucleoprotein xâm nhập vào bộ máy Golgi, nối và gắn protein bề mặt của virus, lắp ráp tạo virion, sau đó ra khỏi bộ máy Golgi, tạo dung hợp với màng sinh chất để ra khỏi tế bào.
19.2.2.1.2. Flaviviridae Thuộc virus Arbo nhóm B gồm các chi: - Flavivirus gồm các virus: + Virus sốt Dengi/ Sốt xuất huyết Dengi. + Virus viêm não Nhật Bản. + Virus viêm não do ve. + Virus viêm não St. Louis. - Pestivirus, gồm: + Virus gây tiêu chảy ở bò. + Virus gây tả lợn. - Virus viêm gan C (HCV). a- Cấu trúc
Hạt virus được bọc bởi vỏ ngoài đường kính 40-60 nm. Vỏ ngoài có các protein M, protein E. - Capsid dạng khối đa diện, cấu tạo từ các protein C. - Lõi là RNA đơn (+), đầu 5’ gắn mũ, đầu 3’ không gắn đuôi nhưng có trình tự không dịch mã. b- Hấp phụ và xâm nhập - Virus gắn gai vào thụ thể glycoprotein bề mặt và xâm nhập theo kiểu dung hợp hoặc nhập bào tạo endosome. - pH thấp trong endosome làm thay đổi cấu hình trong protein E dẫn đến dung hợp với vỏ ngoài với màng endosome đẩy nucleocapsid vào tế bào chất. - Việc cở vỏ còn chưa rõ. c- Tổng hợp protein Bản đồ RNA genome gồm: 5’.mũ-UTR-C-prM-E-NS1-NS2a-NS2b-NS3-NS4a-NS4b-NS5-3’UTR-đuôi - NS1 có trong vỏ ngoài và trong lumen - Phức hợp protein không cấu trúc NS2b-NS3 là protease dùng để cắt các protein khác. - NS4a và NS3 có chức năng chưa rõ. - Peptidase tín hiệu cắt các protein còn lại. - NS5 là RNA polymerase phụ thuộc RNA. d- Sao chép genome - NS3 và NS5 hoạt động cùng nhau trong phiên mã. - Cả RF và RI đều tham gia vào phiên mã rRNA. Ở đây RF - dạng sao chép là sợi đôi trung gian được tổng hợp từ sợi đơn; RI - dạng trung gian sao chép là một số RNA được tổng hợp cùng lúc trên cùng 1 sợi khuôn genome. - RNA virus (vRNA tức là genome) được tổng hợp nhiều gấp 10 lần c-RNA (RNA bổ sung). - vRNA là khuôn để tổng hợp c-RNA. - mRNA gắn mũ và đuôi, để tổng hợp protein virus. - RNA cũng dùng làm genome.
e- Lắp ráp và giải phóng - Tiến hành dịch mã tạo protein vỏ ngoài, gắn vào màng mạng lưới nội chất và tạo protein cấu trúc. - RNA (có mũ và đuôi) trước hết tổng hợp sợi RNA (-) tạo sợi RNA kép dùng để phiên mã tạo RNA genome. - Genome cuộn lại, gắn với protein C tạo capsid vào màng lưới nội chất theo kiểu nhập bào tại nơi đã gắn protein vỏ ngoài. - Protein pr.M tạo phức với protein E để ngăn cản dung hợp với màng trong. - Protein pr.M gắn với protein C để bắt đầu nảy chồi vào màng lưới nội chất. - Các hạt virus được bọc trong bọng ra khỏi mạng lưới nội chất rồi dung hợp với màng tế bào để ra ngoài. - pr.M cắt thành M để virus có thể nhiễm vào tế bào mới.
19.2.2.1.3. Picornaviridae Gồm các chi: - Enterovirus: + Polio gây bại liệt. + Coxsackia Avà B gây viêm màng não, viêm họng rộp, bệnh Bornholm. + Echo gây viêm màng não. + EV71 gây bệnh tay chân miệng. - Cardiovirus (ECM) gây viêm não, cơ tim ở gặm nhấm. - Rhinovirus gây bệnh đường hô hấp trên, cảm lạnh thường. - Apthovirus gây bệnh lở mồm long móng ở động vật có móng chẽ. - Hepatovirus gây viêm gan A (HAV). - Parechovirus typ 1 và 2 gây bệnh ở người (HPEV 1 & 2). a- Cấu trúc
Virion có kích thước nhỏ (20-30 nm). Capsid dạng khối đa diện, không có vỏ ngoài, chứa các protein VP1, VP2, VP3 nằm trên mặt virion, VP4 nằm bên trong liên kết với RNA. Lõi chứa genome RNA đơn, (+) cuộn chặt trong capsid. Ở virus bại liệt đầu 5’ liên kết với peptide Vpg (thay cho mũ), đầu 3’ gắn đuôi poly (A). Phần mã hoá của genome được chia làm 3 phần P1, P2 và P3. P1 mã hoá cho protein cấu trúc VP1, VP2, VP3 và VP4. P2 mã hoá cho các protein 2A, 2B và 2C. 2A là protease ngăn cản dịch mã của tế bào chủ. 2B và 2C cần cho sự sao chép, trong đó 2C gắn với RNA, có hoạt tính ATP-ase và GTP-ase. Polymerase chính của virus là C3Dpro tham gia phân cắt polyprotein nhưng có điểm cắt khác 2A. b- Hấp phụ và xâm nhập - Virus gắn vào thụ thể CD155 trên bề mặt tế bào, vị trí bám là một “hẽm” (canyon) trên bề mặt capsid. - Virus xâm nhập theo cơ chế thực bào, tạo endosome. - Bơm proton trong endosome tạo pH khoảng 5, gây biến tính protein capsid, làm thay đổi cấu hình, lộ ra acid amin kỵ nước (không phân cực). - Các acid amin này tương tác với lớp lipid của màng endosome, giải phóng RNA vào tế bào chất. c- Phiên mã, sao chép và dịch mã - Phiên mã và sao chép là cùng 1 quá trình và sử dụng enzyme như nhau. - VPg có chức năng mồi trong sao chép. - RNA genome có chức năng mRNA, tham gia dịch mã tạo polyprotein. Polyprotein lại được protease phân cắt thành các phân tử đơn lẻ, có chức năng khác nhau. - Protein 3Dpol là RNA polymerase tiến hành tổng hợp sợi DNA (-). Sợi này sau đó được dùng làm khuôn để tổng hợp genome. - Nằm trước vùng mã hoá ở đầu 5’ là cấu trúc bậc 2 IRES là “bến đỗ của riboxom” nhờ vậy mà quá trình dịch mã không cần mũ. d- Lắp ráp và giải phóng - Trước hết tạo protome 5S gồm VP0, VP1 và VP3. - 5 protome tạo pentame 12-14S. 12 pentame tạp procapsid 73S, chứa 60 protome. - RNA kết hợp với vỏ capsid tạo provirion 155S. - Khi VP0 phân cắt thành VP2 và VP4 thì provirion sẽ trở thành virion hoàn chỉnh 155S. - Virus được giải phóng do tan bào, đó là hệ quả của việc virus tạo enzyme ức chế các quá trình phiên mã, dịch mã của tế bào. Virus được phóng thích tiếp tục lây nhiễm vào các tế bào khác. 19.2.2.1.4. Retroviridae Gồm các chi: (1) Avian-leukosis-sarcoma - Virus sarcoma Rous (RSV) gây ung thư ở gia cầm. - Virus gây bệnh nguyên hồng cầu ở chim (AEV – avian erythroblasttosis virus). - Virus gây bệnh bạch cầu tuỷ bào (MC - myelocytomatosis). (2) Mammalian type-C - Virus gây ung thư bạch cầu chuột Moloney (Mo-MLV-Moloney murine leukema). (3) Virus typ B - Virus gây ung thư vú chuột (MMTV – Mouse mammary tumor virus). (4) Virus typ D - Virus nhiễm ở khỉ Mason-Pfizer (MPMV – Mason – Pfizer Monkey virus). (5) Virus gây ung thư bạch cầu tế bào T ở người (HTLV – Human T-cell leukemia virus) hoặc ở bò (BLV – borine leukemia virus). (6) Lentivirus - Virus gây suy giảm miễn dịch ở người (HIV-1, HIV-2), Visna/Msedi. (7) Spumavirus - Virus tạo bọt ở người. a- Cấu trúc
- Vỏ ngoài có nguồn gốc từ màng sinh chất với các protein: + Protein gai gp 120. + Protein gai gp 41. Hai glycoprotein này gắn với nhau nhờ liên kết S – S để tạo gp 160. + Protein vỏ ngoài P17 – P18. - Capsid dạng khối trụ, chứa protein capsid P24 – P25. - Lõi: + Genome là 2 sợi RNA đơn. (+) giống nhau. + Chứa protein nucleocapsid P9 - P7 gắn quanh genome. + Chứa enzyme phiên mã ngược (RT), integrase, Protease. b- Hấp phụ và xâm nhập của HIV - Protein gai gp 120 gắn vào thụ thể dặc hiệu trên bề mặt tế bào, ví dụ CD4 của tế bào T. - Tiến hành dung hợp thông qua trung gian là vùng kị nước của protein màng TM. - Tuỳ thuộc vào loại virus và tế bào chủ, sự dung hợp cũng xảy ra sau khi virus vào tế bào theo con đường nhập bào. - Virus cởi vỏ và nucleocapsid vào tế bào chất. c- Phiên mã ngược - Phiên mã ngược xảy ra bên trong nucleoprotein trong tế bào chất - Sơ đồ RNA (+) Mũ-R-U5-PBS-gag-pol-env-U3-R-AAAA - mRNA có mũ và đuôi poly A. - 1 phân tử tRNA của tế bào đặc hiệu cho mỗi loại virus gắn vào trình tự tương bù tại vị trí gắn mồi (PBS) và tiến hành phiên mã ngược tạo cDNA. - Enzyme RT hoạt động như một DNA polymerase phụ thuộc RNA và cũng có hoạt tính ribonuclease H. + Bước nhảy 1: Trình tự R của cDNA nhảy sang bắt cặp với trình tự R của RNA và tổng hợp cDNA. + Ribonuclease H phân huỷ toàn bộ genome RNA cũ chỉ còn để lại một mẫu ngắn (trình tự P+) tại vùng env để làm mồi. + Enzyme RT bắt đầu tổng hợp sợi DNA (+) về phía U5 để tổng hợp U3 R U5. + Bước nhảy 2: đoạn U3-R-U5-PBS nhảy để PBS của DNA (+) bắt cặp với PBS của DNA (-). + Cả 2 sợi đều được kéo dài tạo phân tử DNA kép provirus với 2 đầu là 2 đoạn lặp đảo chiều LTR (long terminal repeats) gồm U3-R-U5. d- Gắn vào nhiễm sắc thể - Protein nền matrix có vai trò vận chuyển DNA vào nhân. - Provirus trong nhân khép vòng. - Intergrase tạo điểm đứt sole trong DNA tế bào chủ tại trình tự nhận diện provirus (att). - DNA virus được cài xen vào DNA vật chủ tạo phân tử lai. - Các base không bắt cặp sẽ bị loại bỏ và một đoạn khuyết ngắn sẽ được lấp đầy nhờ DNA polymerase. Kết quả là loại ra 4 cặp base khỏi DNA provirus. e- Sao chép genome và dịch mã - Một khi được cài xen, genome virus nằm dưới sự kiểm soát của tế bào chủ và được phiên mã nhờ RNA polymerase II của tế bào. - Đoạn U3 chứa promoter và enhancer. Phiên mã (cũng là sao chép) bắt đầu từ vị trí +1 của trình tự R tạo bản sao mRNA 35S giống với genome virus, được gắn mũ và đuôi poly (A), vừa dùng để dịch mã tạo protein dùng làm genome. - Nhờ dịch khung (frameshift) mà tạo thành poly protein gag-pol. - Protease của virus phân cắt polyprotein thành các protein cấu trúc và không cấu trúc riêng lẻ. - Sự cắt nối tạo mRNA 24S, gắn mũ, đuôi để mã hoá cho protein env (protein vỏ ngoài). - Gen gag được ưu tiên dịch mã nên protein cấu trúc vượt trội protein không cấu trúc. Các protein tích luỹ trong tế bào chất trong đó 1 số bị cắt trong quá trình chế biến sau lắp ráp. f- Lắp ráp và giải phóng - Các thành phần khác nhau của virus liên kết với nhau cả trước khi polyprotein bị cắt thành các protein riêng lẻ. - Hai sợi RNA (+) liên kết cộng hoá trị với nhau tại đầu 5’ và gắn với nucleoprotein. - Protein capsid gắn với protein nền M và với nucleoprotein. - Protein vỏ ngoài cài sẵn vào màng sinh chất. Nucleocapsid nảy chồi ra ngoài.
19.2.2.1.5. Togaviridae Gồm các chi: - Alphavirus có các virus viêm não ngựa Miền Tây, viêm não ngựa Miền Đông, viêm não ngựa Venezuela, virus rừng Semliki, virus Sindbis và gây nhiễm ở động vật không xương sống. - Rubivirus: virus Rubella (sởi Đức). - Pestivirus: virus gây tiêu chảy bò, tả lợn. - Arterivirus: virus viêm động mạch ngựa.
a- Cấu trúc
- Virus có kích thước trung bình (50-70nm). Vỏ ngoài rất dày với các gai cấu tạo từ các protein E1, E2, E3. + Capsid: dạng khối đa diện cấu tạo từ các protein C. Đầu C hình thành giá đỡ cứng ở phía ngoài của capsid lõi. + Lõi: RNA đơn, (+) đầu 3’ và 5’ có các đoạn tương bù nên bắt cặp với nhau. b- Hấp phụ và xâm nhập - Gai glycoprotein gắn vào protein thụ thể bề mặt của tế bào mà bản chất còn chưa rõ. Dùng kháng thể gắn E2 thì virus không vào tế bào được. - Virus vào tế bào theo cách nhập bào, tạo endosome. - Endosome dung hợp với lysosom. pH thấp trong endosome làm thay đổi cấu hình protein E1 và E2 dẫn đến dung hợp màng, giúp virus thoát ra tế bào chất. - Riboxom gắn trên protein C dẫn đến cởi vỏ. c- Phiên mã và dịch mã sớm
- Dịch mã từ bộ 3 AUG tạo protein p1234, là do đọc mã vượt qua codon dừng. - Enzyme cắt protein p1234 để được 4 protein: + NSp1 có hoạt tính polymerase cho cRNA và RNA mũ của virus. + NSP2 là protein dùng cắt polypeptide, có hoạt tính polymerase cRNA và tổng hợp RNA 26S dưới genome. + NSP3 là protein được phosphoryl hoá, chưa rõ chức năng. + NSP4 là RNA polymerase phụ thuộc RNA, protein 123 cắt thành 3 protein, trong đó protein thứ 2 là protease dùng để cắt polypeptide. d- Sao chép genome - Dịch mã tạo protein NS (không cấu trúc). - Protein NS hình thành phức hợp với RNA polymerase để tạo RNA (-) từ sợi RNA kép trung gian, tiến hành phiên mã tạo vRNA và mRNA (gắn mũ và đuôi) để tổng hợp protein. - Phiên mã mRNA 26S e- Phiên mã và dịch mã muộn - Từ v-RNA phiên mã tạo sợi RNA kép trung gian để phiên mã tạo mRNA, gắn mũ và đuôi. - Dịch mã tạo các protein E1, P62,E2 và3), gắn vào màng mạng lưới nội chất, E1, P62, E2 và E3 kết hợp với nhau tạo gai bề mặt dạng trime. f- Lắp ráp và giải phóng: - Protein cấu trúc (C) gắn với vRNA để tạo provirion, xâm nhập vào mạng lưới nội chất, theo kiểu thực bào, sau đó hoàn thiện tạo virion, ra khỏi mạng lưới nội chất tạo bọng rồi ra khỏi tế bào nhờ dung hợp màng bọng với màng sinh chất. 19.2.2.2. Virus RNA đơn, âm Ba họ đại diện là Rhabdoviridae, Orthomyxoviridae và Paramyxoviridae. 19.2.2.2.1- Rhabdoviridae Có 2 chi là: (1) Vesiculovirus gây bệnh chốc mép và ở động vật không xương sống. (2) Lyssavirus gây bệnh dại, ngoài ra virus này cũng gây bệnh ở thực vật. a- Cấu trúc
Virus có hình viên đạn, kích thước 70x170nm. - Vỏ ngoài là lipoprotein. Trên bề mặt có protein G, tạo gai glycoprotein. Bên trong vỏ ngoài là lớp protein nền (M). - Nucleocapsid xoắn, gồm các protein: Protein nucleocapsid (N) gắn với genome, Photphoprotein (P), protein L. - Lõi chứa RNA đơn, (-) với đầu lặp đảo chiều cho phép bắt cặp với nhau tạo cấu trúc cán chảo. - RNA polymerase phụ thuộc RNA gắn vào cấu trúc lõi. b- Hấp phụ và xâm nhập - Virus gắn protein G vào thụ thể bề mặt, chưa rõ bản chất, của tế bào, có thể là photohatidyl serin. - Khi gắn không cần năng lượng nhưng khi xâm nhập cần 37oC. Nếu ở 10oC thì không xâm nhập được. - Virus vào theo lối nhập bào tạo endosome. Endosome dung hợp với lysosom. - pH thấp trong endosome cảm ứng để vỏ ngoài virus dung hợp với màng endosome và giải phóng nucleocapsid vào tế bào chất. - Phiên mã và sao chép genome tiến hành trong cấu trúc nucleocapsid lõi. c- Phiên mã mRNA - Trong tế bào chất gRNA kháng enzyme ribonuclease khi tạo phức với protein N.
- Protein L cùng với protein P và các yếu tố của tế bào tạo phức hợp gắn vào đầu 3’ phức hợp tự phosphoryl hoá. - Phiên mã bắt đầu từ đầu 3’ và kết thúc sau khi gắn đuôi poly (A) vào gen đầu tiên, tạo ra nhiều loại mRNA để tổng hợp protein N, P, M, G và L. - Tổng hợp protein: + Từ các mRNA tương ứng tổng hợp các protein N, P, M, G, L. + Protein G cài vào màng mạng lưới nội chất, tạo bọng rồi mang ra gắn vào màng tế bào chất. + Protein M sắp xếp sát màng tế bào và gắn với protein G. d- Sao chép genome - Khi lượng protein N tăng lên sẽ phong bế vị trí khởi đầu phiên mã trên genome (vRNA). - Sợi mRNA có chiều dài đủ (bằng genome) được tổng hợp nhờ phức hợp polymerase. + Các yếu tố của tế bào chủ có vai trò quan trọng trong sao chép genome virus. + Lượng protein P tăng lên là cần thiết đối với quá trình sao chép. + Hầu hết gRNA nhanh chóng tạo cấu trúc ribonucleoprotein. Một số gRNA dùng để phiên mã mRNA. e- Lắp ráp và giải phóng - Phức hợp ribonucleoprotein lỏng lẻo liên kết với protein M, ngăn cản sự sao chép và phiên mã của virus. - Khi có nhiều protein M hơn, nó sẽ gắn vào ribonucleoprotein xoắn chặt, ngăn cản sự phiên mã và đưa chúng tới màng sinh chất để nảy chồi ra ngoài. 19.2.2.2.2. Orthomyxoviridae Gồm 1 chi Orthomyxovirrus với các virus cúm A, B, C, cúm gia cầm H5N1,... a- Cấu trúc
Virus đa hình thái, đường kính 80-120nm - Vỏ ngoài có các protein sau: + Protein heamagglutinin (HA) hay gai H, gồm 2 tiểu đơn vị HA1 và HA2. Virus cúm A có 16 gai H. + Protein neuraminidase (NA) hay gai N. Virus cúm A có 9 loại gai N. + Protein kênh ion M2. + Protein nền M1. - Capsid dạng xoắn gồm phức hợp ribonucleoprotein (RNP), chứa các protein: Protein nucleocapsid (NP), protein polymerase (PA), protein polymerase (PB1), protein polymerase (PB2). Genome là RNA đơn, âm, phân đoạn. Cúm A và B gồm 8 đoạn, cúm C có 7 đoạn RNA. b- Hấp thụ và xâm nhập - Gai H bám vào thụ thể là acid sialic trên màng tế bào, rồi xâm nhập vào tế bào theo lối nhập bào, tạo endosome rồi dung hợp với lysosom. - pH thấp trong endosome giúp protein dung hợp (protein F) nằm ẩn phía trong gai H chồi lên, cắm vào màng endosome đẻ đẩy nucleocapsid vào tế bào chất. Enzyme từ lysosom cũng có thể phân giải màng endosome. - Nucleocapsid vào nhân, tiến hành phiên mã và sao chép trong nhân. c- Phiên mã Phức hợp polymerase gồm PA, PB1 và PB2. - RNA virus gắn vào các vị trí gắn đầu 3’ và 5’ trên PB1, RNA của tế bào gắn vào PB2. - Virus chiếm đoạt mũ ở đầu 5’ của mRNA của tế bào để làm mồi cho mRNA của mình nhờ enzyme exonuclease, vì thế nên mới phải chui vào nhân. - Tạo mRNA rồi ra khỏi nhân. d- Tổng hợp protein - Tiến hành tổng hợp các loại protein của virus ở ngoài tế bào chất sau đó chui vào nhân để tạo nucleocapsid. e- Sao chép genome Khi protein NP tích luỹ nhiều, chúng sẽ bám và phong bế mũ ở đầu 5’ và các gốc 4-7 A trên RNA của virus, ngăn chặn sự lặp lại của các gốc U của vRNA và tạo thành cRNA (tức sợi RNA + bổ sung). - cDNA được protein NP bao quanh dẫn đến việc phức hợp polymerase mất đi protein PA và tiến hành sao chép vRNA từ khuôn cRNA mới tổng hợp. - vRNA được bao bởi protein Np để tạo nucleocapsid. f- Lắp ráp và giải phóng - Phức hợp RNP hình thành trong nhân, cùng với protein M1 được khuếch tán thụ động vào nhân đồng thời cũng có sự gắn protein bề mặt virus vào màng sinh chất của tế bào. - M1 gắn với RNP dẫn đến vận chuyển ra khỏi nhân. - Protein NA cắt gốc acid sialic để giúp virus nảy chồi thoát khỏi tế bào. 19.2.2.2.3. Paramyxovirida Gồm các chi: - Paramyxovirus gồm virus Sendai (á cúm typ 1 ở chuột), á cúm typ 1và 3 ở người. - Rubelavirus gồm virus quai bị, Newcastle, á cúm typ 2, 4ª, 4b ở người. Có protein HN. - Morbillivirus gồm sởi, dịch sốt chó (canine distemper). Có gai H nhưng không có gai N. - Pneumovirus, virus hợp bào hô hấp ở người (HRSV – Human respiratory syncytial virus). Không có gai H và N. a- Cấu trúc
Virus có kích thước 125-250 nm. - Vỏ ngoài gồm các protein: + Protein F (dung hợp). + Protein heamagglutinin – neuraminidase (HN) có hoạt tính gắn và cắt acid nucleic. + Protein nền (M) nằm ngay dưới vỏ ngoài . - Nucleocapsid dạng xoắn gồm: + Genome là một phân tử RNA đơn, (-). + Protein nucleocapsid (NP) bao quanh genome, liên kết với protein M và các protein L và P. + Protein lớn (L) có hoạt tính polymerase. + Photphoprotein (P). b- Hấp thụ và xâm nhập - Virus gắn gai HN vào thụ thể bề mặt là acid sialic của tế bào ở pH trung tính. - Protein F được protease của tế bào chủ cắt và cho phép dung hợp với màng tế bào để đưa nucleocapsid vào tế bào chất. - Protein M gắn vào protein NP, ức chế tổng hợp mRNA và điều này phải được thực hiện trước khi phiên mã sớm. c- Phiên mã mRNA - Các gen trên gRNA tách biệt nhau nhờ trình tự gắn nằm giữa các gen gọi tắt là ICS (short intercistronic nucleotide sequences). Đây là RNA đa gen (polycistron).
- Các sản phẩm của các gen chồng lớp và tiếp đó là các sản phẩm của các gen P/C/V/D dùng trong điều hoà chu trình nhân lên. - Phiên mã tiến hành trong cấu trúc RNP lõi, bắt đầu tại đầu 3’ và kết thúc khi thêm đuôi poly (A) vào gen đầu tiên. - Số lượng bản sao mRNA nhiều hay ít tuỳ thuộc vào sự tái khởi động phiên mã. Phiên mã hết gen này rồi đến gen khác rồi quay lại tái khởi đầu phiên mã để được các protein NP, P, M, F, HN và L. d- Tổng hợp protein - Tiến hành tổng hợp protein từ mRNA trong tế bào chất. - Protein F và Hn cài vào màng mạng lưới nội chất, hình thành bọng rồi chuyển đến gắn vào màng tế bào. - Protein M liên kết với protein gai trong màng tế bào chất. - Protein NP phong bế vị trí khỏi đầu và ICS cho phép bắt đầu phiên mã sợi cRNA (RNA tương bù). e- Sao chép genome - Từ sợi gRNA (-) làm khuôn tổng hợp sợi tương bù cRNA (+), rồi đến lượt cRNA mới sinh ra làm khuôn để tổng hợp genome RNA. - gRNA mới sinh lại làm khuôn để tổng hợp mRNA và thêm nhiều cRNA. - gRNA liên kết với các protein NP, P và L để tạo cấu trúc lõi. f- Lắp ráp và giải phóng - Protein NP gắn vào vị trí đặc hiệu bao quanh RNA tạo nucleocapsid nảy chồi ra ngoài. - Các protein heamagglutinin – neuraminidase (HN) cắt acid sialic nằm trên bề mặt tế bào cho phép virus thoát khỏi tế bào. 19.2.2.2.4- Reoviridae Bao gồm các chi: - Orthoreovirus. Virus reo typ 1 ở người. - Rotavirus. Virus rota gây bệnh tiêu chảy ở trẻ em. - Orbivirus. Virus gây bệnh lưỡi xanh ở gia súc, virus gây bẹnh sốt Colorado do ve - Phytoreovirus. Virus bệnh lúa lùn, virus khối u ở thực vật. a- Cấu trúc
Virus có đường kích trung bình (60-80nm) không có vỏ ngoài, capsid gồm 2 lớp vỏ: - Vỏ capsid ngoài cấu tạo gồm các capsome và các tiểu đơi vị: d1 – protein hấp phụ. m1 và m16 – protein capsid ngoài. d3 – protein capsid ngoài. - Vỏ capsid trong chứa các protein: l1 – protein capsid trong (nằm ở bề mặt trong của capsid trong). a2 – protein capsid trong. m2 – protein lõi. a4 – protein không cấu trúc. m3 – protein không cấu trúc. l3 – RNA polymerase. - Genome gồm 10-11 phân tử RNA kép: L1, L2 và L3: RNA kép dài. M1, M2 và M3: RNA kép trung bình. S1, S2, S3 và S4: RNA kép ngắn. b- Hấp phụ và xâm nhập - Gai VP4 của virus bám vào thụ thể bề mặt glycophorin – A của tế bào lông ruột. - Virus vào tế bào theo có chế nhập bào qua trung gian thụ thể, tạo endosome. - Màng endosome dung hợp với màng lysosom. Enzyme lysosom phân giải vỏ capsid ngoài, giữ lại vỏ capsid trong và tìm cách thoát vào tế bào chất. c- Phiên mã và sao chép - RNA polymerase phụ thuộc RNA do virus mang theo được hoạt hoá khi vỏ ngoài bị phân huỷ bởi enzyme protease. - RNA tiến hành phiên mã tạo mRNA trên khuôn sợi RNA (-). Quá trình thực hiện bên trong vỏ capsid. - mRNA có 2 chức năng: mRNA và RNA khuôn để tổng hợp sợi tương bù cho genome. - mRNA được đẩy qua kênh nằm ở đỉnh capsid vào tế bào chất. d- Dịch mã - mRNA tổng hợp trên riboxom các loại protein cấu trúc và không cấu trúc (enzyme). - Protein cấu trúc lắp ráp tạo vỏ capsid trong, bao quanh 11 đoạn RNA (+). Tiếp đó sợi RNA (+) đuợc dùng làm khuôn tạo sợi RNA (-) tương bù để được genome RNA kép. e- Lắp ráp và giải phóng - Protein cấu trúc lắp ráp tạo vỏ capsid ngoài bao quanh capsid trong, tạo virion hoàn chỉnh, phá vỡ tế bào ra ngoài. Tế bào lông ruột bị phá huỷ, gây tiêu chảy. 19.2.3. Bacteriophage Bacteriophage (Bacterriophage), viết tắt phage (phage), là virus kí sinh ở vi khuẩn. Genome của chúng có thể là DNA hoặc RNA, với kích thước nằm trong khoảng từ 2,5 đến 150kb. Phage có thể có chu trình sống đơn giản – chu trình tan hoặc phức tạp – chu trình tiềm tan, ở đó genome của chúng được cài vào NST của tế bào hoặc hoạt động theo phương thức chuyển vị. Phage được phát hiện từ đầu thế kỷ XX, một cách độc lập, bởi hai nhà khoa học là Twort (1915) và d’Herelle (1917). Phage được nghiên cứu rất sâu rộng như là một mô hình về virus và như một công cụ để phát hiện những kiến thức cơ bản đầu tiên về DNA (DNA là vật chất di truyền, xác định mật mã di truyền, sự tồn tại của mRNA và nhiều khía cạnh sinh học phân tử cơ bản khác).
Vì phage kí sinh ở prokaryota nên chúng thường có trình tự quan trọng giống như tế bào chủ. Do vậy, chúng còn được sử dụng như là mô hình đơn giản cho nhiều khía cạnh sinh học phân tử khác nhau của prokaryota. Phage được sử dụng phổ biến như là vectơ tách dòng, liệu pháp gen, sản xuất vacxin. Nhiều enzim do phage mã hóa được dùng trong kỹ thuật di truyền để nối các đoạn gen và biểu hiện gen trong tế bào prokaryota…
Các quá tình nhân lên ở bacteriophage Về cơ bản, các bước nhân lên của các bacteriophage trong tế bào vi khuẩn là giống nhau. Tuy nhiên, cũng có những khác biệt ở từng loại. Một số nhân lên theo chu trình tan, số khác lại nhân lên theo chu trình tiềm tan. a. Hấp phụ: Trong môi trường dịch thể, phage ở trạng thái chuyển động tự do. Do va chạm ngẫu nhiên, phân tử protein ở đầu mút sợi lông đuôi gắn đặc hiệu vào thụ thể - phân tử polysaccharit, trên bề mặt màng ngoài của E.coli. Mỗi loại phage gắn vào một loại thụ thể, có thể là polysaccharit trên bề mặt tế bào Gram (-) hoặc acid teichoic của vi khuẩn Gram (+), số khác chỉ gắn được vào đầu mút của pili F. b. Xâm nhập: Phage tiết lyzozim phá hủy peptidoglycan. Bao đuôi co lại, ống trục đâm xuyên qua thành tế bào, đẩy acid nucleic vào trong tế bào. c. Tổng hợp các thành phần: Sau khi acid nucleic vào tế bào là thời kỳ ẩn (eclipse period). Không có bất kỳ một virion nguyên vẹn nào được tạo thành. Genome của virus kiểm soát bộ máy tổng hợp của tế bào, ngăn chặn sự tổng hợp bình thường của tế bào để chuyển sang tổng hợp các thành phần của phage bao gồm tổng hợp genome và protein (y như kẻ xâm lược trong một quốc gia không phòng thủ). d. Lắp ráp: Các bộ phận đầu, đuôi, lông, v.v. được tạo thành ở các nơi khác nhau như trong phân xưởng, sau đó lắp ráp ngẫu nhiên với nhau. Genome DNA được tạo thành nhờ DNA polymerase mới tổng hợp sau đó chui vào lõi để tạo virion hoàn chỉnh. e. Phóng thích: Lyzozim do phage tổng hợp phá hủy peptidoglycan thành tế bào. Khi thành tế bào bị phá hủy, sự thẩm thấu sẽ làm cho tế bào trương lên và vỡ ra. Phage giải phóng ra môi trường xung quanh để lặp lại chu trình nhân lên ở tế bào mới. Sau đây là một số phage có tầm quan trọng trong thực tiễn. (1) Phage M13 Phage có dạng sợi, chứa genome DNA (+) đơn, khép vòng, với kích thước 6,4Kb. M13 gắn đặc hiệu vào pili F (pili được mã hóa bởi plasmit, gọi là yếu tố F, chỉ có ở tế bào “đực”) của E.coli thông qua protein phụ (g3p) nằm trên vỏ ở đầu sợi phage. Sự gắn này cảm ứng làm thay đổi cấu trúc của protein chính tạo nên vỏ capsid khiến toàn hạt co ngắn lại tạo động lực bơm DNA vào tế bào chất. DNA polymease của tế bào tổng hợp một sợi bổ sung tạo DNA kép dạng sao chép (RF). Genome gồm 10 gen nằm sát nhau, có một vùng xen nhỏ (intergenic region) ở đó chứa điểm khởi đầu sao chép (ori). Phiên mã xảy ra (vẫn nhờ enzyme của tế bào chủ) tại bất kỳ một trong số vài promoter, cho đến khi gặp một trong hai điểm kết thúc (terminator). Quá trình này dẫn đến những gen nằm gần điểm kết thúc được phiên mã nhiều hơn so với các gen nằm xa, và đây cũng là phương thức chủ yếu trong việc điều hòa sự biểu hiện gen của phage M13. Tất cả RF đều được tổng hợp như tổng hợp DNA kép bình thường, nhưng sự khởi đầu sao chép cần phải cắt đứt sợi (+) nhờ enzyme endonuclease (sản phẩm của gen 2) để hở đầu 3’-OH, mà không cần mồi. Từ mỗi RF liên tục tạo ra các sợi DNA (+) dành cho đóng gói tạo virion. Các sợi này không dùng được làm khuôn tạo sợi DNA (-) vì ngay sau khi được tổng hợp, chúng đã được bao bọc bởi protein, sản phẩm của gen 5. Cấu trúc này được chuyển ra màng tế bào, ở đó DNA bám vào protein chính của capsid trên màng tế bào rồi chui ra theo lối nảy chồi mà không phá vỡ tế bào. Vi khuẩn nhiễm M13 vẫn tiếp tục sinh trưởng và phân chia, cho dù với tốc độ thấp hơn, tạo ra thế hệ tế bào mới và giải phóng M13. Lượng DNA được chui là ra khác nhau, nên các hạt có kích thước khác nhau. Trong quần thể luôn có những hạt mang nhiều genome và những hạt mang một phần genome. Nhờ cách nhân lên kỳ lạ và cấu tạo genome đặc biệt mà M13 được sử dụng có hiệu quả trong công nghệ sinh học: - Genome là chuỗi đơn nên được dùng trong giải trình tự. - Dạng trung gian RF (DNA kép khép vòng) như là một plasmit nên được dùng làm vectơ tách dòng. - Không có sự hạn chế về kích thước genome nên có thể gắn một đoạn DNA ngoại lai lớn. - Do không làm tan tế bào, nên có thể duy trì và liên tục tách chiết một lượng lớn DNA tách dòng. (2) Phage T7 Cũng như các phage khác, T7 cũng có đầu hình khối đa diện gắn với một đuôi ngắn với 6 sợi lông đuôi. Phage T7 có kích thước nhỏ, có genome là DNA kép (khoảng 40 Kb) dạng thẳng. Trước hết đầu mút sợi lông đuôi gắn vào thụ thể LPS trên màng sinh chất. Khi xâm nhập, một đầu của genome (gọi là đầu trái) bao giờ cũng đi vào trước, do vậy bản đồ genome được chia theo phần trăm tính từ đầu trái. Ngay sau khi genome vào tế bào chất, RNApolymease phụ thuộc DNA bình thường của E.coli nhận ra 3 promoter của các gen sớm ngay, nằm giữa vị trí 1% và 2%, ở đầu 3’ của genome. Tất cả các sản phẩm của các gen sớm đều tham gia vào sự điều khiển làm dừng phiên mã của tế bào chủ, chuẩn bị cho sao chép và biểu hiện genome virus. Bản phiên mã sớm được ribonuclease III của tế bào chủ phân cắt thành các mRNA, trong đó có mRNA mã hóa cho protein kinase (gp0,7) dùng để phosphoryl hóa và làm bất hoạt RNA polymerase của vật chủ, ngăn cản sự phiên mã của các gen sớm khác. Và tổng hợp một RNA polymerase (gp1) dùng để phiên mã các gen nằm ở giữa, tham gia vào sao chép DNA T7 và các gen muộn tham gia vào lắp ráp và làm tan bào để giải phóng virus. Các sản phẩm khác, gp2, cũng gắn vào RNA polymerase của tế bào để làm bất hoạt enzyme này. RNA polymerase của T7 có tính đặc hiệu cao với promoter T7, nên ức chế RNA polymerase của tế bào và dành toàn bộ hoạt động phiên mã cho T7. Sự sao chép genome T7 gồm 3 giai đoạn: a. Khởi đầu diễn ra tại một vị trí nằm ở đầu trái của genome. RNA polymerase (gp1) của T7 tổng hợp một mồi ngắn từ các protomer tại vùng khởi đầu dùng cho bước khởi đầu. b. Kéo dài từ mồi theo 2 hướng, do 2 enzyme gp5 và gp7 của T7 xúc tác. Protein gp5 gắn với thioredoxin để tạo một DNA polymerase dùng cho tổng hợp DNA từ mồi, theo hướng 5’ – 3’, do đó tạo thành sợi muộn. Protein gp4 là enzyme đa chức năng, có hoạt tính pimase, helicase và NTP-ase, nó gắn vào mạch DNA đơn duỗi xoắn của phân tử DNA mẹ và chuyển dịch theo hướng 5’ – 3’ của DNA nhờ thuỷ phân NTP. Sự mở xoắn của chuỗi DNA mẹ theo hướng chạc sao chép. Khi gặp vị trí đặc hiệu nhận biết primase, gp4 sẽ tổng hợp các mồi tetraribonucleotide (pppApCpCpC hoặc pppApCpCpA). Các mồi này cung cấp đầu 3’-OH để khởi đầu tổng hợp sợi muộn nhờ DNA polymerase của T7. Protein gp3 là một endonuclease và gp6 là một exonuclease tiến hành phân giải DNA vật chủ để cung cấp các nucleotide làm nguyên liệu cho tổng hợp DNA T7. Gp6 và DNA polymerase I của vật chủ tham gia vào loại bỏ mồi mRNA. c. Sự tạo thành phân tử DNA trùng lặp (concatemer) Do phân tử RNA mồi ở đầu 5’ bị loại bỏ trước khi hoàn thành sao chép, nên mỗi phân tử DNA kép mới tạo thành, có một đoạn ở đầu 5’ không được sao chép và tương ứng với nó ở đầu 3’ của mạch tương bù là đoạn đơn. Các đoạn đơn này của hai chuỗi có trình tự lặp lại cùng chiều, nên có thể gắn với nhau, tạo phân tử DNA đúp (bimolecule). Những chỗ hở được tổng hợp bổ sung và nối với nhau nhờ ligase. Concateme được endonuclease T7 (gp3) cắt tại các vị trí đặc hiệu ở cả hai phía tạo thành các đoạn đầu sole, sau đó DNA polymerase hoàn tất đầu mạch đơn (sole) thành phân tử kép đầu bằng, có kích thước của genome T7. Phage trưởng thành thoát khỏi tế bào nhờ lyzozim (gp3,5) làm tan bào. Đáng chú ý là RNA polymerase rất đặc hiệu với promoter T7, cho nên có thể tiến hành phiên mã bất kỳ trình tự DNA nào nếu có promoter T7. RNA polymerase của T7 có khả năng phiên mã mạnh gấp 5 lần RNA polymerase của E. coli, do đó thường được dùng để biểu hiện gen trong E. coli. (3) Phage T4 T4 là một phage T chẵn, có kích thước lớn. Genome là DNA kép, dạng thẳng, chứa khoảng 100 gen với kích thước 1,7x108 bp. Chu trình nhân lên bắt đầu khi lông đuôi bám vào thụ thể bề mặt của E. coli, tiết lyzozim phân giải peptidoglycan thành tế bào, rồi bơm DNA vào trong tế bào. Mặc dù genome là dạng thẳng nhưng trong quần thể, các virion chứa genome có trình tự thay đổi theo kiểu vòng tròn, tức là đổi đầu quay vòng (circular permutation) và có hai đầu lặp lại cùng chiều. Do vậy để dễ hình dung, người ta thường biểu diễn bản đồ gen của T4 là một vòng tròn. Ví dụ các virion có thể chứa các genome có trình tự sau đây: ABCDEFGAB, CDEFGABCD, EFGABCDEF, GABCDEFGA, … Một trong những gen được dịch mã đầu tiên là enzyme dùng để chặt nhỏ DNA của tế bào chủ, nhưng không động chạm đến DNA của phage, bởi vì base cytosin thông thường trong DNA đã được thay thế bởi một base C cải biến, 5-hydroxy metylcytosin (HMC). HMC bắt cặp với guanin nên không làm thay đổi chức năng khuôn của virus, nhưng sự có mặt của nó giúp cho enzyme virus phân biệt được acid nucleic của virus với của tế bào chủ. Sự cải biến này làm cho DNA của phage kháng lại nhiều enzyme giới hạn của tế bào chủ. Tuy nhiên, các nhóm hydroxyl của 5-HMC được cải biến bằng cách thêm gốc glucozơ, DNA đã glucosyl hoá này kháng lại hầu như tất cả các endonuclease của tế bào chủ. Toàn bộ cho trình từ lúc bắt đầu tiếp xúc với bề mặt tế bào đến khi làm tan tế bào kéo dài 20-30 phút ở 3oC, trong đó tổng hợp DNA bắt đầu khoảng 5 phút sau khi nhiễm. Genome T4 gồm 3 nhóm gen sớm ngay, gen sớm sau và gen muộn. - Gen sớm ngay được phiên mã nhờ RNA polymerase (bình thường aabb’ và d) đã được cải biến của tế bào để tạo các protein điều hoà. Khi xâm nhập, một protein (sản phẩm của gen alt (alternation) của T4) được đưa vào tế bào chủ cùng với DNA để cải biến trình tự RNA polymerase E. coli, làm giảm ái lực của yếu tố d với enzyme lõi (aabb’) khiến yếu tố này không nhận diện được promoter của E. coli dẫn đến cản trở phiên mã của tế bào, nhưng lại nhận diện được promoter gen sớm của T4 để tiến hành phiên mã. - Gen sớm sau mã hoá cho enzyme DNA polymerase tham gia và sao chép và phiên mã của virus. - Gen muộn mã hoá cho các protein cấu trúc (đầu, đuôi, lông) và enzyme dùng để giải phóng phage khỏi tế bào. Sự sao chép DNA diễn ra tương tự như ở phage T7. Phân tử mới được sao chép có đầu thừa tận cùng tại đầu 5’ của cả hai phân tử con. RNA mồi không được thay thế bởi DNA, do đó ở đầu 3’ đối diện là đoạn DNA đơn. Các base bổ sung ở đầu 3’ của 2 phân tử trùng lặp (concateme). DNA ligase hàn các chỗ đứt. Enzyme giới hạn cắt phân tử DNA concateme thành các đoạn bằng nhau tạo genome đổi đầu quay vòng. Phiên mã genome muộn hoàn toàn phụ thuộc vào sự có mặt của DNA mới tổng hợp, là dạng có cấu trúc lỏng lẻo thuận lợi cho phiên mã. Ba peptide, sản phẩm của các gen T4 (33,45 và 55) gắn vào RNA polymerase với các chức năng là các chất hoạt hoá gen (gen activators) giúp nhận diện promoter gen muộn. Quá trình lắp ráp và giải phóng diễn ra như ở các phage T khác. Tế bào bị phá vỡ, giải phóng 100-200 hạt phage. DNA-ligase của T4 là enzyme xúc tác cho phản ứng tạo liên kết photphodieste giữa 3’-OH và 5’-P liền kề của phân tử DNA và RNA sợi đôi. Do enzyme này có khả năng nối các đoạn DNA có đầu sole hoặc đầu bằng, nên được sử dụng rộng rãi trong tách dòng và cải biến DNA. (4) Phage lamda Phage lamda là virus ôn hoà có cấu tạo giống T4, nhưng đuôi của nó chỉ có một sợi lông ngắn. Đầu chứa DNA kép, dạng thẳng. Sự lây nhiễm bắt đầu khi sợi lông đuôi gắn vào thụ thể bề mặt của E. coli, sau đó tiêm DNA vào tế bào chất. DNA của viron là sợi kép, dạng thẳng nhưng 2 đầu là mạch đơn và bổ sung cho nhau, gọi là đầu dính (cohesive end), viết tắ là cos. Ở trong tế bào chủ, hai đầu genome nhanh chóng gắn vào nhau tạo phân tử dạng vòng. Điều gì xảy ra tiếp theo là tuỳ thuộc phương thức sinh sản, chu trình tan hay chu trình tiềm tan. Chu trình tan về cơ bản giống với T4, nhưng ở đây chia làm giai đoạn. - Giai đoạn I: Từ điểm khởi đầu Ori, tiến hành sao chép theo hai hướng đối diện nhau, theo kiểu theta và kết thúc khi 2 chạc ba gặp nhau, để tạo nhiều khuôn cho phiên mã và khuôn cho sao chép. - Giai đoạn II: Từ khuôn DNA kép dạng vòng mới được tổng hợp, tiến hành sao chép theo cơ chế vòng tròn xoay (giống như fx174). Enzyme cắt làm đứt sợi (+) ngoài để lộ đầu 3’-OH, sợi (-) nguyên vẹn bên trong dùng làm khuôn. Sợi (-) xoay, trong khi các nucleotide gắn vào đầu 3’-OH để nối dài mạch mà không cần mồi, thường sau khi tổng hợp một đoạn DNA mạch đơn dài sẽ gắn mồi để tổng hợp mạch bổ sung, tạo phân tử DNA kép. Enzyme cắt do phage l mã hoá sẽ cắt ở vị trí chuyên biệt trên cả hai mạch để tạo đoạn có kích thước genome. Điểm cắt là đầu dính cos. Sau sao chép genome là phiên mã gen muộn để tạo protein cấu trúc, tiếp dó là các giai đoạn lắp ráp và giải phóng. Điều đáng chú ý là trên DNA của l có một đoạn (nằm giữa gen J và gen att) không đóng vai trò quan trọng trong quá trình nhân lên của l. Do đó trong kỹ thuật di truyền, phage l được dùng làm vectơ chuyển gen. Người ta cắt bỏ đoạn này và thay vào đó đoạn gen mong muốn để tách dòng. Chu trình tiềm tan. Ngược với chu trình tan là chu trình tiềm tan, theo đó không một thành phần nào của phage được tổng hợp và tế bào không bị tan. Genome khép vòng được cài xem vào NST của tế bào. DNA ở trạng thái này gọi là prophage. Một protein ức chế do phage mã hoá ngăn cản sự phiên mã của các gen muộn, do đó duy trì trạng thái tiềm tan. Khi tế bào phân chia, prophage cũng được phân chia theo. Trạng thái này sẽ bị mất khi chất ức chế bị bất hoạt. Ở trạng thái prophage, một vài gen của phage (ngoài gen mã hoá cho chất ức chế) vẫn có thể được biểu hiện, đôi khi làm thay đổi phenotyp của tế bào chủ, ví dụ làm thay đổi hình dạng khuẩn lạc. Các ví dụ đáng chú khác là độc tố của vi khuẩn Corynebacterium diphteriae, gây bệnh bạch cầu, được mã hoá bởi prophage. Độc tố thần kinh do Clostridium botulinum có trong các hộp thịt phồng, cũng do prophage mã hoá. Tế bào tiềm tan có thể sinh trưởng, phân chia và hình thành tế bào mới giống như tế bào mẹ. Trong thực tế, hầu hết trong nuôi cấy, vi khuẩn đều có thể mang một hoặc vài prophage, nhưng chỉ một tỷ lệ rất nhỏ 1/10000 hoặc 1/100000 prophage có thể tách khỏi nhiễm sắc thể để tiến hành chu trình tan, tạo virion mới và gải phóng ra môi trường. Các virion được giải phóng không ảnh hướng tới các tế bào tiềm tan khác, bởi vì protein ức chế luôn duy trì prophage ở trạng thái tiềm tan, giúp tế bào tiềm tan “miễn dịch” trước sự xâm nhập của virus cùng loại. Do có sự “miễn dịch” nên các phage ôn hoà chỉ tạo vệt tan khi cho xâm nhiễm vào các chủng không phải tiềm tan. Các vệt tan xuất hiện rất đặc trưng. Các virus độc tạo vệt tan rất rõ trong khi virus tiềm tan chỉ tạo vệt tan mờ. Các vệt tan nhìn thấy được vì nhiều tế bào bị tan do các phage tiến hành chu trình tan. Vệt tan mờ là vì một số tê bào tiềm tan vẫn sống sót, sinh sản trong vệt tan. (5) Phage chuyển vị (TP) Phage chuyển vị (transposable phage) chủ yếu gặp ở các vi khuẩn Gram âm, nhất là Pseudomonas. Một trong các ví dụ điển hình nhất là phage Mu. Phage chuyển vị có vòng đời theo cả chu trình tan và tiềm tan, giống như phage l, nhưng klhác ở phương thức sao chép genome. Ở pha tan sớm, một số quá trình phức tạp liên quan đến enzyme transposase của virus, DNA polymerase của vi khuẩn và một số enzyme khác (của virus và vi khuẩn) tiến hành sao chép DNA và cài bản sao genome mới tạo thành của virus vào tế một vị trí nào đó trên NST của tế bào và do đó có thể làm bất hoạt các gen tại vị trí gắn dẫn đến đột biến. Không có genome nguyên ven của virus rời khỏi NST tế bào chủ. Chỉ sau một số vòng chuyển vị sao chép (replicative transposition), genome virus cùng với một phần DNA liền kề của tế bào được tách ra khỏi tế bào để thực hiện chu trình tan. Phage chuyển vị được dùng phổ biến trong công nghệ sinh học để đưa gen mong muốn vào các tế bào nhân sơ.
19.3. NUÔI CẤY VIRUS ĐỘNG VẬT 19.3.1. Phát triển phương pháp nuôi cấy Trước kia nếu muốn nuôi cấy virus thì người ta phải nuôi cấy trên cơ thể động vật. Như vậy rất khó quan sát được các hiệu ứng đặc thù của virus ở mức độ tế bào. Vào những năm 30 của thế kỷ 20 các nhà virus học đã phát hiện thấy trứng gà mang phôi (đã thụ tinh hay chưa thụ tinh) có thể được dùng để nuôi cấy virus herpes, virus đậu mùa và virus cúm. Mặc dù phôi gà có kết cấu giản đơn hơn nhiều so với cơ thể thỏ hay chuột thí nghiệm nhưng phôi gà vẫn là một cơ thể phức tạp. Sử dụng phôi gà không giải quyết được triệt để những vấn đề gặp trong hiệu ứng tế bào học do virus gây ra. Mặt khác, vi khuẩn cũng phát triển tốt trên phôi gà, trên các phôi nhiễm vi khuẩn rất khó đánh giá chính xác được các hiệu ứng do virus. Vì vậy ngành virus học đã phát triển chậm chạp khi chưa cải tiến được phương pháp nuôi cấy virus. Có hai phát hiện mới đã làm triển khai được phương pháp nuôi cấy tế bào giúp cho các nhà virus học và các nhà khoa học khác. Một là, việc phát hiện và sử dụng chất kháng sinh để ngăn ngừa sự cảm nhiễm vi khuẩn. Hai là, các nhà sinh học phát hiện thấy các enzyme thủy phân protein (proteolytic), nhất là trypsin, có thể làm tách riêng các tế bào ra khỏi các mô xung quanh mà không làm tổn hại đến các tế bào này. Rửa các tế bào và đếm số lượng chúng, pha loãng rồi chuyển vào các bình, các ống nghiệm hay hộp Petri bằng nhựa (plastic). Các tế bào trong dịch huyền phù sẽ hấp phụ trên bề mặt lớp thành nhựa, cúng sinh sôi nẩy nở và lan ra thành một tầng tế bào gọi là tầng tế bào đơn (monolayer). Tầng tế bào đơn này có thể truyền thế hệ (subcultured). Nuôi cấy truyền thế hệ (subculturing) là đem các tế bào nuôi cấy được đi nuôi cấy trong các môi trường nuôi cấy mới. Một lượng lớn các tế bào truyền thế hệ (subcultures) được tạo thành từ một mẫu tổ chức đơn sẽ là mẫu tế bào nuôi cấy đồng nhất cần thiết cho hàng loạt các nghiên cứu hiệu ứng của virus. Thuật ngữ nuôi cấy mô (tissue culture) đã được sử dụng rộng rãi nhưng nuôi cấy tế bào (cell culture) cần sử dụng chính xác hơn. Hiện nay, phần lớn các tế bào nuôi cấy đều là tầng tế bào đơn sinh trưởng ra từ các tế bào phân tán đã được xử lý nhờ enzyme. Nhờ sử dụng nhiều loại hình nuôi cấy tế bào vận dụng chất kháng sinh để ức chế cam nhiễm mà Virus học đã bước vào thời đại vàng (Golden Age). Từ thập kỷ 50 đến thập kỷ 60 của thế kỷ 20 đã có trên 400 virus được phân lập và giám định (characterized) Mặc dù các virus mới đã được phát hiện nhưng trọng tâm hiện nay là cân giám định kỹ hơn các đặc tính của virus, xác định các bước cảm nhiễm (infection) và nhân lên (replication) của virus. Các thí nghiệm vệt tan (plaque) dùng trong nghiên cứu phage (phages) có thểt dùng cho cả các virus đồng vật. Chẳng hạn, nuôi cấy tầng tế bào đơn các tế bào người, sau đó cấy virus, nếu virus làm tan tế bào thì rất nhiều vòng xâm nhiễm sẽ sản sinh vệt tan. 19.3.2. Các loại hình nuôi cấy tế bào Trong Virus học lâm sàng và Virus học thực nghiệm thường sử dụng rộng rãi 3 kiểu cơ bản nuôi cấy tế bào: -Nuôi cấy tế bào nguyên đại (primery cell culture) -Các chủng nguyên bào sợi nhị bội thể (diploid fibroblast strains) -Các dòng tế bào liên tục (continous cell lines) Vật nuôi cấy tế bào nguyên đại là lấy trực tiếp từ động vật, không qua truyền thế hệ (not subcultured). Các động vật lấy mẫu càng non thì thời gian sống của tế bào nuôi cấy càng kéo dài. Điển hình của các tế bào này là thể hỗn hợp của nhiều loại hình tế bào như tế bào cơ bắp (muscle), tế bào thượng bì (epithelial). Tuy vậy số lần phân cắt của các tế bào này thường không nhiều, nhưng cũng đủ để hỗ trợ cho nhiều chủng virus sinh trưởng Nếu việc nuôi cấy tế bào nguyên đại có thể truyền thế hệ một cách lặp lại thì một típ (type) tế bào sẽ chiếm ưu thế và vật nuôi cấy các tế bào này được gọi là chủng tế bào (cell strain). Trong chủng tế bào tất cả các tế bào loại hình gene giống nhau. Chúng có thể truyền thế hệ nhiều lần, mỗi thế hệ giữa các tế bào khả năng xuất hiện các khác biệt là rất nhỏ. Các khác biệt này có thể can thiệp vào kết quả xác định hiệu ứng của virus. Các chủng nguyên bào sợi nhị bội thể là các chủng tế bào thường dùng nhất. Nguyên bào sợi (fibroblasts) là một loại tế bào chưa thành thục. Chúng có thể sản sinh ra collagen và các sợi khác như là cơ chất của mô liên kết giống như chân bì (dermis of the skin) Các chủng tế bào này là lấy từ mô của phôi, giữ tốc độ phát triển nhanh và phân cắt liên tiếp của phôi. Chủng tế bào này hỗ trợ cho sự phát triển của các loại virus và thường không mang các virus ô nhiễm như các chủng tế bào thu được từ các động vật trưởng thành. Chính vì vậy mà các chủng tế bào này thường được sử dụng để sản xuất các loại vaccin virus. Típ thứ ba nuôi cấy tế bào được sử dụng rộng rãi là dòng tế bào liên tục. Một dòng tế bào liên tục chứa các tế bào có thể sinh sản ra rất nhiều thế hệ. Dòng tế bào liên tục nổi tiếng nhất là dòng tế bào HeLa. Dòng tế bào này sau khi được phát hiện từ năm 1951 đã được nuôi cấy liên tục và được các nhà nghiên cứu khắp thế giới sử dụng rộng rãi. Nguồn gốc của dòng tế bào HeLa là lấy từ một phụ nữ bị ung thư tử cung và chữ HeLa là lấy từ các chữ đầu của tên người này. Trên thực tế rất nhiều dòng tế bào liên tục thời kỳ đầu đều là dùng các tế bào ung thư ác tính, vì chúng có năng lực sinh trưởng rất nhanh. Các dòng tế bào vĩnh sinh (immortal cell lines) được nuôi cấy trong các phòng thí nghiệm không bị lão hóa, phân chia nhanh chóng, nhu cầu dinh dưỡng thường đơn giản hơn các tế bào thông thường. Ví dụ dòng tế bào HeLa có chứa 2 gen virus cần thiết cho năng lực vĩnh sinh hóa (immortality) của chúng. Dòng tế bào vĩnh sinh này là dị bội thể (heteroploid) –chứa số lượng khác nhau các nhiễm sắc thể, vì vậy mà có tính đa dạng di truyền. Việc nuôi cấy tế bào đã thay thế trên mức độ rất lớn các động vật thực nghiệm và trứng chứa phôi khi nghiên cứu Virus học động vật. Tuy vậy trứng gà mang phôi vẫn là hệ thống vật chủ tốt nhất đối với virus cúm A.
Ngoài ra chuột nhỏ loại Albino Swiss vẫn được dùng để nuôi cấy Arbovirrus (Arthropod-borne viruses), có lúc cũng dùng những dòng tế bào động vật có vú khác hoặc các dòng tế bào muỗi. 19.3.3. Hiệu ứng bệnh biến tế bào (The cytopathic effect) Hiệu ứng thấy được của virus đối với tế bào cảm nhiễm được gọi là hiệu ứng bệnh biến (cytopathic effect, CPE). Các tế bào trong nuôi cấy biểu hiện các loại hiệu ứng chung, bao gồm sự biến đổi hình dạng tế bào và sự tách ra khỏi các tế bào phụ cận hoặc vật dụng nuôi cấy.
Tuy nhiên CPE có thể cũng có những biểu hiện đặc biệt mà các nhà nghiên cứu virus học thực nghiệm có thể dùng để giám định sơ bộ virus xâm nhiễm. Chẳng hạn, adeno virus và herpes virus ở người có thể làm cho tế bào cảm nhiễm trương phồng lên do bị tích lũy dịch thể. Còn picorna virus lúc xâm nhập tế bào có thể ngăn chặn tế bào và làm tan vỡ tế bào khi chúng thoát ra. Paramyxo virus có thể làm dung hợp (fuse) các tế bào đứng gần nhau, tạo nên các thể hợp bào (syncytia, số nhiều là syncytium). Thể hợp bào có thể chứa từ 4 tới 100 nhân trong một tế bào chất chung. Ngoài ra có một dạng CPE khác là sự biến nạp (transformation): từ tế bào bình thường biến thành tế bào ác tính. 19.4. VIRUS VÀ UNG THƯ Ung thư được biết là loại bệnh mà có sự rối loạn về hoạt động và chức năng của những tế bào bình thường. Có thể định nghĩa Ung thư (cancer) như là sự tăng trưởng không thể khống chế được của các tế bào dị thường, nói cách khác là các tế bào không nhừng phân chia. Rất nhiều trường hợp không thể đình chỉ được sự phân cắt của chúng, kết quả là tạo thành các Mô ung thư (Neoplasm) hoặc Khối u (Tumor). Mô ung thư có thể là lành tính (benign) - tăng trưởng phi ung thư (noncancerous). Nhưng nếu các tế bào xâm nhập và can thiệp vào chức năng của các mô bình thường chung quanh thì sẽ trở thành ác tính (malignant). Ung thư ác tính và các tế bào của chúng có thể di căn (metastasize) hoặc khuếch tán (spread) tới các mô khác của cơ thể. Năm 1911, F. Peyton Rous đã phát hiện thấy một số virus có thể gây ra ung thư ở động vật, ông xác định một số sarcomas (neoplasm ở mô liên kết) ở gà là do một loại virus được gọi là Rous sarcoma virus, RSV- gây ra. Vì vậy không có gì đáng ngạc nhiên với các phát hiện virus có liên quan đến bệnh ung thư ở người. Mặc dù phần lớn ung thư ở người là do các đột biến di truyền (genetic mutation) hay do các hóa chất gây ô nhiễm môi trường, hoặc do cả hai gây ra ung thư. Các nhà nghiên cứu dịch tễ cho rằng có khoảng 15% các ung thư ở người do có sự xâm nhiễm của virus. 19.4.1. Các virus gây ung thư ở người Qua nhiều năm nghiên cứu và thực nghiệm hiện nay đã biết ít ra là đã có 6 loại virus liên quan đến ung thư ở người. Có thể còn có nhiều hơn nữa sẽ được xác định. Virus Epstein-Barr, EBV hay thường gọi là virus EB: là loại virus gây ung thư ở người được hiểu biết nhiều nhất. Loại virus DNA này là một Herpes virus lần đầu tiên phát hiện trên trẻ em Châu Phi bị ung thư lympho Burkitt. Loại ung thư ác tính này gây ra xưng phồng rồi dẫn đến hoại tử hàm dưới. Trên thực tế đã chứng minh được là có 3 loại ung thư khác liên quan đến virus EB. Một số virus Papilloma ở người (human papillomavirus, HPV) có liên quan đến mật thiết đến ung thư ở người. Mặc dù một số virus DNA này chỉ gây ra các mụn cơm (warts) lành tính nhưng có những tip khác (HPV-8, HPV-16) đã gây ra ung thư tử cung (ung thư mô thượng bì). Có thể nói 99,7% ung thư tử cung là do HPV gây nên Một số virus DNA khác gây ung thư là virus viêm gan B (HBV). Chúng gây ra bệnh viêm gan và 80% các trường hợp ung thư gan là do chúng gây nên. Ung thư Sarcoma Kaposi là loại ung thư tế bào nội bì trong mạch máu và hệ thống lympho, loại ung thư này có liên quan đến virus Herpes-8 ở người (human herpesvirus 8). Hiện nay đã phát hiện thấy các virus gây ung thư ở người chủ yếu là các virus hai chuỗi DNA (dsDNA). Tuy nhiên một số virus RNA sợi thẳng, hay(+) sense RNA, nhất là các Retrovirus cũng có thể liên quan đến ung thư. Chẳng hạn HTLV-I gây nên bệnh máu trắng tế bào T ở người lớn hoặc ung thư lympho (aldult T cell leukemia/lymphoma). 19.4.2. Virus ung thư gây bệnh như thế nào? Giống như phage (thực khuẩn thể, phage), các virus động vật thường thông qua việc làm tan để gây chết tế bào. Một số virus động vật khác có thể xâm nhiễm tế bào và tạo thành các Tiền virus (proviruses). Trong một số trường hợp sự xâm nhiễm này có thể làm cải biến về sinh lý và di truyền của các tế bào vật chủ (host cells) - tức là các CPE đã nói tới ở phần trên. Ví dụ các virus RSV đã làm cho các tế bào tách khỏi dụng cụ nuôi cấy và biến thành hình cầu. Trong trường hợp virus ung thư DNA có thể tồn tại dưới dạng tiền virus. Các CPE chủ yếu của chúng là sự phân cắt không kiểm soát được ở các tế bào xâm nhiễm. Quá trình này được gọi là sự biến nạp gây u (neoplastic transformation), là đặc trưng điển hình của các virus DNA. Nhiều virus đã đưa toàn bộ hay một phần DNA xâm nhập vào vị trí ngẫu nhiên trong DNA của vật chủ. Tuy nhiên, chỉ có một ít gen virus là cần thiết cho biến nạp. Virus Papilloma (thuộc họ Papovaviridae) có thể gây ra các tế bào ung thư ở người, nhưng DNA của chúng lại tồn tại tự do trong tế bào chất của vật chủ. Một ít gen của virus Papilloma là có hoạt tính và dẫn đến virus được sao chép (replicate) mỗi khi phân cắt tế bào. Nếu DNA virus ngẫu nhiên kết hợp với DNA trong tế bào vật chủ thì sẽ xảy ra sự tổng hợp protein virus không khống chế được. Các protein này có thể làm cho tế bào mất đi khả năng khống chế phân cắt. Một số protein virus này đã ngăn chặn hiệu ứng của các gen ngăn chặn ung thư (tumor-suppressor genes), các gen có thể ngăn chặn sự phân cắt không khống chế được của tế bào. Không có sản phẩm của các gen này, vật chủ sẽ không khống chế được sự phân cắt tế bào và phát triển thành một khối u. Nhiều virus Retro là các Virus ung thư RNA. Trên thực tế chúng duy nhât là những virus RNA gây ung thư đã biết. Cần nhắc lại, các virus Retro đã dùng enzyme transcriptase ngược để phiên mã sợi RNA (+) vào DNA, sau đó trở nên một tiền virus (provirus) xâm nhập vào nhiễm sắc thể của vật chủ. Protein phiên mã bởi tiền virus HTLV-I sẽ chuyển hóa tế bào vật chủ thành khối u. Sự xâm nhiễm cũng dẫn đến sự phát sinh các hạt virus (virion) mới bằng nẩy chồi, chúng không làm gây chết tế bào bị xâm nhiễm. Do đó, virus ung thư RNA có thể tiếp tục xâm nhiễm vào các tế bào chưa bị xâm nhiễm khác hoặc các tế bào sinh sản (sex cells). Nếu là tế bào sinh sản sự tồn tại của các hạt virus sẽ làm cho thế hệ sau của vật chủ cũng sẽ bị virus xâm nhiễm. 19.4.3. Các gen gây ung thư (Oncogenes) Các protein sinh ra bởi các virus ung thư làm cho các tế bào vật chủ phân chia không khống chế được là được phiên mã từ các đoạn DNA được gọi là các Gen ung thư (oncogenes), theo tiếng Hy Lạp thì onco là tích thành khối. Trong các virus DNA gây ung thư, các gen ung thư không chỉ gây ra khôi u mà chúng còn chứa các thông tin để tổng hợp protein virus cần thiết cho sự sao chép virus. Gen gây ung thư ở virus RNA gây ung thư thì hoàn toàn khác. Các nhà virus học và tế bào học đều đã phát hiện thấy một số virus RNA gây ung thư có thể sinh ra các gen đặc biệt (extra genes) từ các tế bào vật chủ bình thường khi sao chép virus. Các gen này rất giống với gen gây ung thư và được gọi là gen Nguyên u (Proto-oncogenes). Một gen Nguyên u là một gen bình thường, sau khi bị virus khống chế nó có thể dẫn đến việc tế bào mất khả nămg khống chế sự phân chia, tức là nó biến thành gen gây ung thư. Nhiều gen gây ung thư đã được tìm thấy trong các virus gây ung thư và phần lớn mang thông tin di truyền có thể làm cho tế bào phân chia không hạn chế. Các gen gây ung thư này là các gen đột biến chứa sự mất đoạn (deletions) hoặc sự thay thế (substitution). Các đột biến này gây ra sự biến đổi cấu trúc trong protein mà gen đã phiên mã. Gen gây ung thư hoạt động theo một trong hai phương thức sau đây: Một là, sản phẩm của gen gây ung thư có thể gây ra sự gián đoạn chức năng của tế bào bình thường dẫn đến sự phân cắt tế bào. Hai là, gen gây ung thư bị khống chế bởi các gen diều hòa của virus (viral regulators) nằm gần vị trí gắn kết (intergration) chúng vào nhiễm sắc thể của vật chủ, gen điều hòa này ”mở ra” (”turn on”) gen tạo ra các protein bình thường, nhưng là sự tổng hợp quá mức hoặc là tổng hợp không phù hợp với chu kỳ sống của tế bào vật chủ; ngoài ra còn xuất hiện sự phân chía quá mức tế bào. Sự phát hiện ra gen gây ung thư ở virus có ý nghĩa quan trọng trong việc giúp hiểu biết về ung thư. Về ung thư ở người còn rất nhiều vấn đề cần tiếp tục nghiên cứu, nhưng các thuốc đặc hiệu chống virus trong tương lai có thể ngăn ngừa được các bệnh ung thư do virus gây nên. 19.4.4. Hy vọng phòng chống ung thư Ung thư là một trong những nguyên nhân chính gây tử vong trên thế giới. Năm 2005, người ta ước tính có khoảng 7,6 triệu nạn nhân chết vì bệnh này và trong mười năm sắp tới, số người chết sẽ là 84 triệu nếu như không phòng chống. Tổ chức Y tế thế giới (WHO: World Health Organization) đã thống kê trong 20 năm qua số người mới mắc ung thư hàng năm đã tăng từ 10 triệu lên đến 15 triệu. Số người chết vì ung thư mỗi năm đã tăng từ 6 triệu lên đến 10 triệu. Phòng chống ung thư là một sự nghiệp hết sức to lớn đối với cả thế giới. WHO đã đề nghị mục tiêu toàn cầu là làm giảm mức tử vong xuống 2% mỗi năm, từ 2006 đến 2015 để có thể tránh hơn 8 triệu người chết trong số 84 triệu người sẽ chết vì ung thư dự tính cho mười năm sắp tới và WHO cố gắng đạt mục tiêu này. Những nước nghèo có hơn 70% số người chết do bệnh ung thư vì thiếu tiền để chẩn đoán lẫn trị bệnh. Chỉ mỗi chứng nhiễm độc thuốc lá mà cũng đã gây khoảng 1,5 triệu cái chết mỗi năm. WHO nỗ lực tích cực. Người ta ước tính có khoảng 40% bệnh ung thư có thể tránh được. Tuy nhiên nguy cơ sẽ tăng rất nhiều khi bệnh nhân mập phì hay bị nhiễm độc thuốc lá. Người ta nhận xét thấy nguy cơ bệnh ung thư và những bệnh mạn tính trên phương diện toàn cầu tăng nhanh khi trong thức ăn hiện nay có quá nhiều mỡ, đường và muối, còn rau quả thì ít đi. WHO họp lần thứ nhất tại Genève từ 6-17/2/2006 để thỏa thuận với nhau về một phương cách giảm tiêu thụ thuốc lá. Các đại biểu cho biết quỹ có 8 triệu USD cho những sinh hoạt chống thuốc lá trong thời gian hai năm. Chiến lược toàn cầu cũng chú trọng việc tránh tiêu dùng các thực phẩm có chứa hóa chất gây ung thư, chương trình chủng ngừa các bệnh nhiễm virus viêm gan trên bình diện quốc tế. Để cải thiện việc phát hiện sớm, điều trị và chăm sóc người bị ung thư, Trung tâm Nghiên cứu Ung thư (IARC: International Agency for Research on Cancer) của WHO đã tìm ra nhuyên nhân gây bệnh ung thư và các cơ chế của sự xuất hiện ung thư đồng thời tìm những phương cách phát hiện sớm. WHO còn kết hợp với những cơ quan ngoài hệ thống Liên hiệp quốc như Cơ quan năng lượng nguyên tử quốc tế, Hội chống ung thư quốc tế và nhiều Viện ung thư quốc gia. Cho đến nay chưa có nổi vaccin chống ung thư ngoại trừ vaccin Gardasil phòng tránh ung thư cổ tử cung. Mục tiêu của vaccin phòng tránh ung thư là sử dụng kháng nguyên ung thư biểu đạt đặc hiệu để kích hoạt, phục hồi và tăng cường các phản ứng miễn dich kháng ung thư, nhằm có thể sát hại, thanh lọc tế bào ung thư. Vì vậy có thể đem vaccin ung thư chia thành 3 loại: loại Dự phòng, loại Chữa trị, và loại vừa Dự phòng và Chữa trị. Cục quản lý Thực phẩm và Thuốc Hoa Kỳ (FDA) đã phê chuẩn vaccin Gardasil để phòng tránh virus HPV- gây ra ung thư cổ tử cung. Vaccin này chống lại 4 nhóm HPV-6, HPV-11, HPV-16, HPV-18. HPV-16 và HPV-18 được coi là thủ phạm của 70% các trường hợp ung thư cổ tử cung, còn HPV-6 và HPV-11 là thủ phạm của 90% các trường hợp mụn cơm ở bộ phận sinh dục (cả nam lẫn nữ). Vaccin này cũng được khẳng định là sẽ giúp phòng ngừa ung thư âm đạo và tử cung do virus HPV. HPV lây truyền qua đường tình dục là một loại virus rất phổ biến. Khoảng 20 triệu người Mỹ bị nhiễm loại virus này, và ở lứa tuổi 50, ít nhất 80% phụ nữ đã từng bị nhiễm các chủng của virus HPV. Không phải phụ nữ nào bị nhiễm virus này cũng sẽ bị ung thư. Tỉ lệ bị bệnh ung thư cổ tử cung do HPV là rất nhỏ.
Virus Human papillomavirus (HPV) thuộc nhóm DNA.
Mặc dù vaccin Gardasil có khả năng bảo vệ tránh khỏi một trong những nguyên nhân hàng đầu dẫn đến căn bệnh ung thư cổ tử cung nhưng vẫn còn có những nguyên nhân khác không phải do virus HPV gây ra căn bệnh ung thư đáng sợ này. Chính vì vậy việc thường xuyên kiểm tra vẫn là rất cần thiết. Những đợt kiểm tra định kỳ sẽ giúp phát hiện ra những thể ung thư khác cũng như nguy cơ có thể bị ung thư do chủng virus HPV nhóm khác. Những nghiên cứu đã cho thấy hiệu quả 100% của vaccin này trong việc chống lại sự lây nhiễm virus HPV-16 và HPV-18 ở người. Các thử nghiệm cho thấy vaccin có hiệu quả tối ưu trong ít nhất 4 năm. Thời gian lâu hơn hiện vẫn chưa được kiểm chứng. Gardasil chỉ chứa cấu trúc phân tử của loại virus này chứ không phải là virus được làm yếu đi như một số loại vaccin khác. Cơ quan FDA Hoa Kỳ thừa nhận Gardasil tốt nhất cho nữ giới trong độ tuổi từ 9-26 và không đưa vào lịch tiêm chủng quốc gia.Công ty Merk & Co có nhiệm vụ tiếp tục nghiên cứu đưa vào sử dụng loại vaccin phòng chống virus HPV gây mụn rộp ở nam giới. Gardasil là thế hệ vaccin đầu tiên phòng ung thư cổ tử cung được cấp phép. Trên thực tế, đây là loại vaccin đầu tiên chống lại một nhân tố gây ung thư. Tháng 4/2007 vaccin chống ung thư cổ tử cung Cervarix cũng xin phép được đưa ra sử dụng tại Mỹ
|